ESCAPING FROM PREDATORS An Integrative View of Escape Decisions (2015)
Part II Escape and refuge use: theory and findings for major taxonomic groups
IId Refuge use
9 Hiding time in refuge
José Martín and Pilar López
9.1 Introduction
In most cases, prey animals do not escape randomly, but toward a refuge such as a rock crevice, burrow, tree, or a patch of thick vegetation. Safer habitats where animals restrict most of their activity to avoid exposure to potential predators found in other places are often called refuges or refugia (e.g., Werner et al. 1983). That is not our meaning of refuge in this chapter. Here, a refuge refers to small portions of the habitat where a prey hides temporarily after escaping from a predator that has been detected or that has attacked the prey. By definition, a refuge is a physical structure or some part of a habitat where the predator cannot enter to follow and capture the prey, or is at least impeded from doing so. Therefore predation risk immediately decreases, sometimes to zero, when the prey enters a refuge and hides inside it.
Numerous studies have shown that prey often respond to predator presence by increasing their use of refuges or safe microhabitats (Lima & Dill 1990; Sih et al. 1992). However, most prey animals do not spend their lives in the safety of a refuge (Sih 1997) because by staying in refuges prey often incur some fitness costs that should be minimized (e.g., the loss of time available for foraging; Godin & Sproul 1988; Koivula et al. 1995; Dill & Fraser 1997; Martín et al. 2003a; Blumstein & Pelletier 2005; or mate searching; Sih et al. 1990; Martín et al. 2003b; Reaney 2007). Unfavorable conditions in refuges (e.g., suboptimal temperatures or oxygen levels) might also entail physiological costs, such as hypothermia or hypoxia (Wolf & Kramer 1987; Martín & López 1999b; Polo et al. 2005). And, finally, refuges are often useful against some types of predators, but can expose of prey to other type of predators, which can cause a conflict in refuge use (Soluk 1993; Sih et al. 1998; Amo et al. 2005).
9.2 The optimal hiding time
Because refuge use is often costly, prey should optimize the time spent hiding inside a refuge and elect to use alternative escape strategies if refuge costs are too high. Some theoretical models have analyzed when prey should decide to leave the refuge and resume their behavior after a predator’s unsuccessful attack (see Chapter 2 for details). The hiding decision should be made based on the balance between the cost of being outside the refuge (i.e., predation risk) and the costs of refuge use. This decision is very important, as suggested by a state-dependent dynamic model that explored potential fitness consequences of hiding decisions in yellow-bellied marmots (Marmota flaviventris; Rhoades & Blumstein 2007). This model suggested that the overall survival of a population is substantially reduced when individuals make suboptimal hiding decisions (e.g., hiding 50% less or 200% more than the optimal hiding time substantially decreases the likelihood of survival). Because individuals need simultaneously to acquire energy and avoid predation, individuals making incorrect decisions could either starve or be killed.
Economic models of refuge use predict that prey should choose to leave a refuge when the cost of refuge use is higher than the risk to fitness of predation outside (Martín & López 1999a; Cooper & Frederick 2007; see Chapter 2 for details and graphs). The optimal hiding time will be modified in two ways. First, when the rate of reduction of predation risk decreases, the time spent in the refuge should increase. Second, when the costs of refuge use increase, the time spent in the refuge should decrease.
Economic models make three predictions. (1) Time spent in the refuge should be longer when the threat of the initial attack has been higher, and therefore the subsequent diminution of risk is slower (i.e., the probability of a new attack after emergence is higher). (2) Time spent in the refuge should be longer when costs of refuge use are lower. (3) The effects of costs of refuge use should be more important in the high-risk situation. In the low-risk situation, prey should emerge after a short period of time (when predation risk level drops), regardless of levels of costs of refuge use.
Optimality models consider the costs and benefits of all the possible strategies an animal could adopt, and predict which should be the optimal decision that maximizes fitness. These models predict that the optimal hiding time should increase with initial fitness and decrease as the maximum benefit that can be obtained by not hiding increases (Cooper & Frederick 2007; see Chapter 2 for details and graphs). Initial fitness is important because prey having high residual reproductive value should be more cautious to protect this “reproductive asset” (Clark 1994) by remaining hidden for longer. However, when the benefits obtainable by emerging are greater, prey should accept greater risk, and have shorter hiding times. If large benefits are retainable after death, prey may emerge and allow themselves to be killed. These predictions are similar to those from the previous economic models. However, optimality models make the new predictions. One is that emergence time increases with initial fitness and varies with the exponent in the benefit function. The second prediction, that prey may risk being killed by emerging for large benefits retainable after death, is unique to the optimal hiding time model.
9.3 Experimental tests of the predictions of hiding time models
Models of refuge use make many ordinal level and quantitative predictions about hiding or emergence time based on the balance between the benefits of staying hidden (i.e., risk of emerging) and the costs incurred while hiding in the refuge (i.e., cost of not emerging) (Sih 1992; Martín & López 1999a; Cooper & Frederick 2007).
One example of an experimental test of the predictions from optimal hiding models (see Chapter 2) is a field study that analyzed the variation in hiding time in a refuge (rock crevices) in the Iberian rock-lizard, Iberolacerta cyreni, under two different simulated predation risk levels and under different thermal costs of refuge use (Martín & López 1999a). As predicted by the economic models, lizards hid longer when the risk level of the initial attack was high, and therefore the subsequent reduction of risk with time is slower (Figure 9.1b). However, lizards may reduce hiding time in the refuge while also minimizing predation risk by walking while hidden under rock screes to reappear at a different place, leaving the predator waiting for the prey to appear at the wrong place (Figure 9.1). Interestingly, only individuals with initial high body temperatures seem able to adopt this alternative strategy.
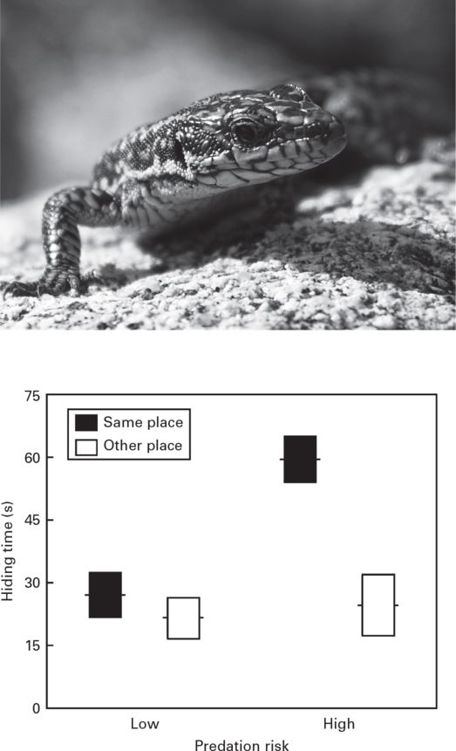
Figure 9.1
(a) Predation risk level affects refuge use by the Iberian rock lizard, Iberolacerta cyreni. (b) Hiding times (mean ± SE) in a refuge after a predatory attack are shown under two levels of risk. Responses change depending on whether lizards emerge from the site of refuge entry or at a different site.
(Figure 9.1a: photograph by J. Martín; Figure 9.1b: redrawn from Martín & López 1999a)
On the other hand, effects of thermal costs are more important in high-risk situations. Under high-risk situations, time spent in the refuge increases when thermal conditions of the refuge are more similar to thermal conditions outside (i.e., when physiological costs of refuge use are lower) (Figure 9.2). Under these high-risk situations, hiding time is correlated with the temperature differential if lizards emerge from the point of refuge entry, but not when lizards emerge from different places. In low-risk situations, the place of emergence has no effect on the relationship between temperature differential and hiding time.
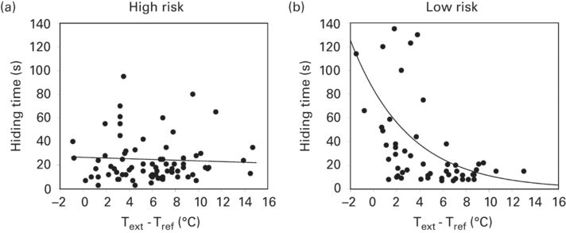
Figure 9.2
Thermal costs affect refuge use by Iberian rock lizards, Iberolacerta cyreni. Relationships observed between hiding time in a refuge after a predatory attack and the difference between the external temperature (Text) and the refuge temperature (Tref) under two levels of risk. Thermal cost of refuge use is higher when this difference is greater because body temperature decreases faster in refuge as refuge temperature decreases relative to the outside temperature.
(Redrawn from Martín & López 1999a)
In addition to this first test of the economic model by Martín and López (1999a) that examined effects of both predation risk and cost of hiding, predictions of models of optimal hiding time have been verified consistently for various specific costs and benefits in several observational and experimental studies (Table 9.1). These are discussed in the remaining sections of this chapter. Of the studies carried out on hiding behavior, there has been a strong bias toward a few species of lizards, with only a minor fraction of studies made with other animals, such as fish, crabs, insects, marmots, etc.
Table 9.1 Studies examining variation in hiding time (or related variables) depending on several factors in different taxa. Results of these studies are useful to test the predictions of models of optimal refuge use.
|
Species |
Factors examined |
Reference |
|
PLANTS: |
||
|
Mimosa pudica |
“Foraging” costs |
Jensen et al. 2011 |
|
INVERTEBRATES: |
||
|
Polychaetes: |
||
|
Balanus glandula |
Foraging costs. Nutritional state. Body size |
Dill & Gillet 1991 |
|
Serpula vermicularis |
Foraging costs |
Dill & Fraser 1997 |
|
Mollusks: |
||
|
Corbicula fluminea |
Physiological costs (oxygen availability) |
Saloom & Duncan 2005 |
|
Margaritifera margaritifera |
Group size (risk dilution) |
Wilson et al. 2003 |
|
Crustaceans: |
||
|
Uca lactea perplexa |
Predators’ risk level. Handling |
Jennions et al. 2003 |
|
Uca mjoebergi |
Foraging and reproductive costs |
Reaney 2007 |
|
Pagurus filholi |
Predators’ risk level. Alternative tactics |
Mima et al. 2003 |
|
Pagurus acadianus |
Foraging costs |
Scarratt & Godin 1992) |
|
Heteroptera: |
||
|
Notonecta maculata |
Predators’ risk level. Foraging costs. Nutritional state. Competition with conspecifics |
Martín & López 2004b |
|
Notonecta hoffmani |
Predator density. Body size. Nutritional state |
Sih 1992 |
|
Gerris remigis |
Reproductive costs |
Sih et al. 1990 |
|
Lepidoptera: |
||
|
Achroia grisella |
Costs of intrasexual competition between males |
Brunel-Pons et al. 2011 |
|
Orthoptera: |
||
|
Gryllus integer |
Prey’s conspicuous calling |
Hedrick 2000 |
|
Long-term differences in predation pressure |
Hedrick & Kortet 2006 |
|
|
Gryllus bimaculatus |
Limb autotomy on calling behavior |
Bateman & Fleming 2006 |
|
Teleogryllus oceanicus |
Prey’s conspicuous calling. Differences in parasitism pressure |
Lewkiewicz & Zuk 2004 |
|
Trichoptera: |
||
|
Halesus radiatus (larvae) |
“Waiting games” between predator and prey |
Johansson & Englund 1995 |
|
Ephemeroptera: |
||
|
Baetis tricaudatus (larvae) |
Multiple types of predators |
Soluk 1993 |
|
Ephemerella subvaria (larvae) |
Multiple types of predators |
Soluk 1993 |
|
Diptera: |
||
|
Culex pipiens (larvae) |
Predators’ risk level. Physiological costs (oxygen availability) |
Rodríguez-Prieto et al. 2006 |
|
VERTEBRATES: |
||
|
Fish: |
||
|
Colisa lalia |
Physiological costs (oxygen availability) |
Wolf & Kramer 1987 |
|
Fundulus diaphanus |
Predators’ risk level. Body size. Nutritional state |
Dowling & Godin 2002 |
|
Foraging costs. Nutritional state |
Metcalfe & Steele 2001 |
|
|
Gasterosteus aculeatus |
Foraging costs |
Godin & Sproul 1988 |
|
Body size. Foraging costs. Body condition |
Krause et al. 1998 |
|
|
Predators’ risk level. Body size. Foraging costs. Nutritional state |
Krause et al. 2000a,b |
|
|
Phoxinus phoxinus |
Predators’ risk level. Body size. Foraging costs. Nutritional state |
Krause et al. 2000a |
|
Poecilia reticulata |
Long-term differences in predation pressure |
Harris et al. 2010 |
|
Semotilus atromaculatus |
Foraging costs |
Gilliam & Fraser 1987 |
|
Frogs: |
||
|
Lithobates catesbeianus |
Predators’ risk level |
Cooper 2011b |
|
Lizards: |
||
|
Plesiodon laticeps |
Predators’ risk level. Multiple attacks |
Cooper 1998 |
|
Eulamprus heatwolei |
Costs of intrasexual competition between males |
Stapley & Keogh 2004 |
|
Pseudemoia entrecasteauxii |
Multiple types of predators |
Stapley 2004 |
|
Sceloporus jarrovii |
Predators’ risk level. Multiple attacks |
Cooper & Avalos 2010 |
|
Sceloporus virgatus |
Thermal costs |
Cooper & Wilson 2008 |
|
Predators’ risk level. Thermal and foraging costs |
Cooper 2009a |
|
|
inter-individual differences (personalities) |
Cooper 2009c |
|
|
Predator’s risk level and starting distance. Thermal costs |
Cooper 2011a |
|
|
Tail autotomy and handling |
Cooper & Wilson 2010 |
|
|
Tropidurus hispidus |
Costs of intrasexual competition between males |
Díaz-Uriarte 1999 |
|
Acanthodactylus erythrurus |
Multiple attacks |
Martín & López 2003a |
|
Lacerta schreiberi |
Thermal costs. Body size |
Martín & López 2010 |
|
Podarcis lilfordi |
Long-term differences in predation pressure |
Cooper et al.2009 |
|
Predator’s risk level. Multiple attacks |
Cooper et al.2010 |
|
|
Podarcis muralis |
Predation pressure and thermal and foraging costs |
Martín & López 1999b |
|
Predator’s risk level. Thermal costs |
Amo et al. 2003 |
|
|
Predator’s risk level. Multiple attacks |
Martín & López 2005 |
|
|
Multiple types of predators |
Amo et al. 2004, 2005 |
|
|
Iberolacerta cyreni |
Predator’s risk level. Thermal costs. Economic model of refuge use |
Martín & López 1999a |
|
Predator’s risk level. Thermal costs. Multiple attacks |
Martín & López 2001 |
|
|
Predator’s risk level. Thermal costs. Multiple attacks |
Polo et al. 2005 |
|
|
Body size related thermal costs |
Martín & López 2003b |
|
|
Thermal and foraging costs. Body condition |
Amo et al. 2007a,b |
|
|
Predator’s risk level. Different individual predators |
Cooper et al. 2003 |
|
|
Foraging costs. Nutritional state |
Martín et al. 2003a |
|
|
Predator’s risk level. Reproductive costs |
Martín et al. 2003b |
|
|
Prey’s conspicuous coloration |
Cabido et al. 2009 |
|
|
inter-individual differences (personalities) |
López et al. 2005 |
|
|
Predator’s risk level. Multiple attacks |
Martín & López 2004a |
|
|
Temporal patterns of risk |
Martín et al. 2009 |
|
|
Predator’s risk level. Monitoring from refuge |
Polo et al. 2011 |
|
|
Iberolacerta monticola |
Prey’s conspicuous coloration |
Cabido et al. 2009 |
|
Turtles: |
||
|
Mauremys leprosa |
Predator’s risk level. Handling. Alternative tactics |
Martín et al. 2005 |
|
Predator’s risk level |
Polo-Cavia et al. 2008 |
|
|
Trachemys scripta |
Predator’s risk level |
Polo-Cavia et al. 2008 |
|
Birds: |
||
|
Poecile montanus |
Foraging costs. Nutritional state |
Koivula et al. 1995 |
|
Mammals: |
||
|
Marmota flaviventris |
Foraging costs. Body size. inter-individual differences (personalities) |
Blumstein & Pelletier 2005 |
|
Predator’s risk level. Foraging costs. Body size. Fitness consequences of hiding decisions |
Rhoades & Blumstein 2007 |
9.3.1 Factors affecting risk of emergence on hiding time
Immediately after a prey hides in a refuge after escaping from a predator, risk of predation (i.e., costs of emergence) outside the refuge is highest; thereafter, risk level decreases gradually as time spent hidden in the refuge increases. This occurs because after some waiting time most predators will leave the area (Hugie 2003). Thus the probability that the predator is waiting for the prey outside the refuge (i.e., probability of a new attack) decreases as hiding time increases. All theoretical models of refuge use predict that hiding time should increase as the initial risk increases or the rate of diminution of predation risk with time decreases (Martín & López 1999a; Cooper & Frederick 2007; see Chapter 2). This has been confirmed for several types of risks in many studies cited below.
9.3.1.1 Risk due to the predator’s behavior
After entering a refuge, prey may assess predation risk level in the exterior from the characteristics of the predator’s behavior during approach, or the predator’s attack behavior before the prey hides. Predation risk may vary in relation to the threat and characteristics of the immediately previous encounter with the predator (Stankowich & Blumstein 2005). The reduction of predation risk might be very fast when the prey has retreated into the refuge as a preventive strategy to elude a detected predator that has not yet attacked. In contrast, in a high predation risk situation (e.g., a direct attack by the predator), the initial level of predation risk is higher, and risk will diminish more slowly with time because the predator is known to pose a greater threat to the prey.
When a simulated predator approaches the prey faster, which indicates higher risk, subsequent hiding times are longer in several species of lizards (broad-headed skinks, Plestiodon laticeps:Cooper 1998; Iberian rock lizards, Iberolacerta cyreni: Cooper et al. 2003; Martín & López 1999a, 2001; wall lizards, Podarcis muralis: Martín & López 2005; striped plateau lizards, Sceloporus virgatus: Cooper 2009a; and Balearic lizards, Podarcis lilfordi: Cooper et al. 2010; Figure 9.3). In contrast, yellow-bellied marmots hide longer in response to a slower approach of the predator (Rhoades & Blumstein 2007), probably because marmots may consider a slow approach more dangerous as many of their predators slowly stalk their prey.
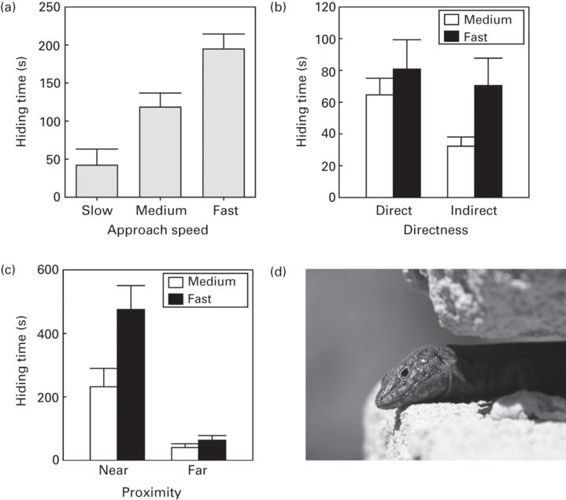
Figure 9.3
Effects of several risk factors on refuge use by Balearic lizards, Podarcis lilfordi (d). Time spent hiding in refuges (mean + SE) by lizards (a) increases linearly with predator approach speed; (b) is greater following direct than indirect approaches that bypass the prey without contact at medium approach speed, but not at the fast approach speed; and (c) is longer when the predator stays near the refuge than farther from it, and the difference between proximity levels of the predator is greater after approaches at the fast than the medium speed.
(Graphs redrawn from Cooper et al. 2010; photograph by J. Martín)
The directness of a predator’s approach is another cue that prey use to assess risk. Predators approaching tangentially might not be attacking, whereas directly approaching predators are more likely to have detected the prey and be preparing to attack. Therefore hiding time is predicted to be longer for direct than indirect approaches. As predicted, when a predator has approached directly, fiddler crabs (Uca lactea perplexa; Jennions et al. 2003) and several lizards (Martín & López 1999a, 2004a, 2005; Cooper et al. 2003, 2010; Cooper 2009a) have longer hiding times than when a predator has approached indirectly (Figure 9.3).
A predator that is closer when first detected or that remains closer to the refuge after the attack may be considered a higher risk leading to longer hiding times in fiddler crabs (Jennions et al. 2003), skinks and other lizards (Cooper 1998, 2009a; Cooper et al. 2010; Figure 9.3). However, in American bullfrogs, Lithobates catesbeianus, time spent submerged after escaping by jumping into water was uncorrelated with proximity of the predator upon escape (Cooper 2011b). This contradiction may be explained because frogs may need to surface to determine whether the predator remains nearby, and can submerge themselves again if so.
Prey responses may also depend on the subtle characteristics of each individual predator, which may be considered to pose different risk levels (Cooper et al. 2003). Also, a prey that was able to escape from a predator that has captured it temporarily may, in a subsequent encounter, assess that this predator, or a similar one, poses a greater risk, and therefore increase hiding time (Cooper & Wilson 2010; Figure 9.4). Duration of handling by the predator before escaping also affected hiding times in fiddler crabs (Jennions et al. 2003) and turtles (Martín et al. 2005).
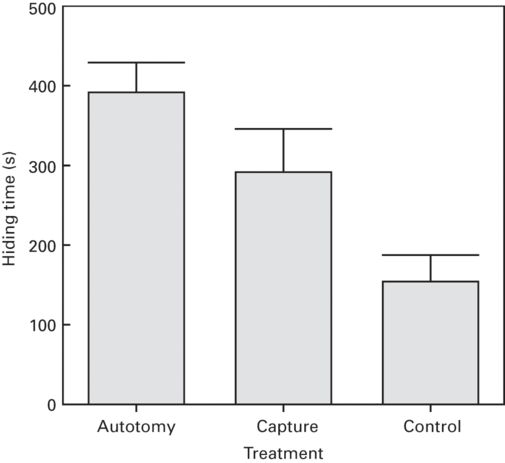
Figure 9.4
Effects of a previous capture by the predator and tail autotomy on refuge use by striped plateau lizards, Sceloporus virgatus. Hiding time (mean + SE) is longer following both tail autotomy and capture than in the uncaptured, unhandled control treatment, and longer following autotomy than just capture and handling.
(Redrawn from Cooper & Wilson 2010)
Multiple repeated approaches also may represent a higher risk, which results in longer hiding times for the second of two consecutive attacks in several lizard species (Cooper 1998, 2011a; Martín & López 2001, 2004a, 2005; Cooper & Avalos 2010; Cooper et al. 2010). Similarly, after the first attack, most individuals of the lizard Acanthodactylus erythrurus do not hide, but stop after running in the open and remain vigilant; whereas in subsequent attacks within a brief interval, lizards use increasingly safer, structurally more complex bush refuges (i.e., larger and with more obstructive cover; Martín & López 2003a). These data suggest that persistent predatory attacks increase predation risk and prey adjust hiding time to correspond with the risk level. However, the outcome of several multiple attacks may be harder to predict than for only two successive attacks because there are usually interactions with other predation risk factors or changes in risk level between attacks (Martín & López 2001, 2004a, 2005; Polo et al. 2005). These combined multiple effects will be examined below in more detail (see section 9.6).
Prey may perceive a higher risk when a given individual predator is persistent. Individual recognition of the predator may not be needed if the assessment is based solely on attack rate. Similarly, prey might respond to an increase in the density of different individual predators (Cooper 1998). Thus aquatic bugs Notonecta hoffmani, increase hiding time in response to increased predator density (Sih 1992). Nevertheless, some prey may be able to use the time interval between successive attacks to estimate whether repeated attacks are originated by the same individual predator, which would indicate persistence of the same individual predator (i.e., high risk), or as independent single attacks by different predators or by the same predator but with low motivation (i.e., low risk) (Martín & López 2004a).
Finally, in contrast to flight initiation distance (see Chapter 2), hiding time seems unaffected by the predator’s starting distance (predator to prey distance when approach begins) in the lizard Sceloporus virgatus (Cooper 2011a), which suggests that once a prey enters refuge, the SD is irrelevant to predation risk upon emergence. However, it is likely that prey dynamically increase their assessed risk such that when there are longer approach times (which should be correlated with SD), there is greater perceived risk (Cooper & Blumstein 2014). Under such conditions, hiding time should also increase as SD increases, just as it increases with increases in other risk factors during the approach.
9.3.1.2 Risk due to the prey’s characteristics
The prey’s own characteristics, such as body size and age, may affect risk perception and refuge use. Small juvenile aquatic bugs Notonecta hoffmani (instar 1) are more susceptible to predation by adults than are large juveniles (instar 3); accordingly, small juveniles show longer hiding times (Sih 1992). Similarly, smaller barnacles, Balanus glandula, have longer hiding times than larger ones (Dill & Gillett 1991). In contrast, large fish often hide longer than small ones (Krause et al. 1998, 2000a,b; Dowling & Godin 2002), possibly because, although at higher risk from predators upon emergence, small fish have higher costs of lost feeding opportunities by hiding than do large fish. Similarly, yellow-bellied marmot pups emerge sooner than adults, which is also consistent with the costs of lost opportunities because marmot pups will not survive hibernation unless they gain sufficient body mass in their first year (Blumstein & Pelletier 2005). It is also possible that young animals must learn about how to respond to risks. In addition, consistent differences in boldness between otherwise similar individuals may affect their risk perception and therefore their refuge use (Blumstein & Pelletier 2005; López et al. 2005; Cooper 2009c; see Chapter 14 for more studies and details).
More conspicuous individuals suffer a greater risk of being detected by predators and therefore should increase hiding time. Male rock lizards, Iberolacerta monticola, have bright-blue UV lateral ocelli that serve as sexual signals, but also make males conspicuous to predators. They seem to compensate for greater conspicuousness by increasing hiding time in proportion to the number of these conspicuous ocelli (Cabido et al. 2009). By contrast, males of a sister lizard species, I. cyreni, that have fewer and less conspicuous ocelli, do not vary hiding time (Cabido et al. 2009). Similar behavioral compensation for increasing conspicuousness of displaying males to predators or parasites has been reported in other animals. For example, male field crickets, Gryllus integer, with longer, more conspicuous, songs behave more cautiously than males with shorter songs; they take longer to emerge from a shelter within a novel, potentially dangerous environment, and they cease calling for a longer time when their calls are interrupted by a predator cue (Hedrick 2000). Similarly, in the field cricket, Teleogryllus oceanicus, latency to resume calling after disturbance corresponds to population-level differences in risk of being parasitized (Lewkiewicz & Zuk 2004). And vividly colored birds are more responsive to the sounds of their aerial predators (Journey et al. 2013).
The ability of the prey to escape again from a predator after emerging from a refuge also influences predation risk and may affect hiding times. For example, lizards that escape by autotomizing their tails subsequently have decreased escape speed and lose the ability to use autotomy again. Thus the risk of being captured upon emergence is greater after an experimentally induced autotomy, which leads to longer hiding times in the lizard Sceloporus virgatus (Cooper & Wilson 2010) (Figure 9.4). Also, field crickets, Gryllus bimaculatus, that have lost legs by autotomy have longer hiding times than intact individuals (Bateman & Fleming 2006).
Finally, the risk dilution effect of being in a group remains almost unexplored in relation to hiding time after a predator attack. However, freshwater pearl mussels, Margaritifera margaritifera, that were in a group closed their valves for shorter times than solitary mussels after experiencing potentially dangerous novel stimuli (Wilson et al. 2012).
9.3.1.3 “Waiting games” between predator and prey
Hiding in a refuge increases safety, but may restrict further information from being obtained about the risk associated with the waiting predator. When this occurs, the individual predator and prey involved become opponents in a ‘‘waiting game” (Hugie 2003). The prey must decide how long to wait for the predator to depart before emerging and potentially exposing itself to attack. The predator must decide how long to wait for the prey to emerge before departing in search of other foraging opportunities. Waiting is assumed to be costly to both players. Hugie (2003) used a game-theoretical model to determine the evolutionarily stable waiting strategy of both players. The model predicts that each player’s waiting distribution (i.e., the distribution of waiting times expected for individuals in each role) will have a characteristic shape: the predator’s distribution should resemble a negative exponential function, whereas the waiting time of the prey is predicted to be more variable and follow a positively skewed distribution. The model also predicts that the predator will rarely outwait the prey.
Illustrating this relationship, Johansson and Englund (1995) studied a waiting game between bullheads, Cottus gobio, which are predatory fish, and casemaking caddis fly larvae, Halesus radiatus. After an unsuccessful attack by a bullhead, a larva remains motionless with its head and legs hidden in a case of organic debris. The bullhead usually responds by orienting toward the larva and remaining motionless. Measures of waiting times of prey and predator reveal a good fit with the predictions of Hugie’s (2003) waiting-game model. Furthermore, as predicted, the predator rarely outwaited the prey: only 1.5% of larvae re-emerged before the bullhead had departed.
9.3.2 Costs of refuge use determining hiding time
When deciding hiding times, prey should consider several costs of refuge use, which are mainly the loss of opportunities for obtaining food or reproductive benefits outside the refuge, and the physiological costs associated with unfavorable conditions inside refuges. The potential for encountering other types of predators inside the refuge is another cost in some situations. Theoretical models predict that the optimal hiding time should decrease as the costs of refuge use increase (Sih 1992; Martín & López 1999a; Cooper & Frederick 2007, see Chapter 2). Several empirical studies have examined how specific costs affect hiding decisions.
9.3.2.1 Loss of foraging opportunities
The cost of remaining hidden in a refuge often varies with food availability outside the refuge, especially when food becomes available in a food-limited environment. Therefore many animals take more risks when food levels are high. For example, the tubeworm, Serpula vermicularis, alters hiding time inside its tube with fluctuating food levels in the environment. It emerges from its tube sooner when food is abundant, thereby avoiding greater opportunity cost of remaining in the tube than when food is scarce (Dill & Fraser 1997; Figure 9.5). Moreover, tubeworms adjust their hiding times in response to short-term experimental changes in food availability. Similarly, juveniles of a small freshwater minnow, Semotilus atromaculatus, spent less time in a safe area that protected them from predators as food level increased outside in the riskier environment (Gilliam & Fraser 1987).
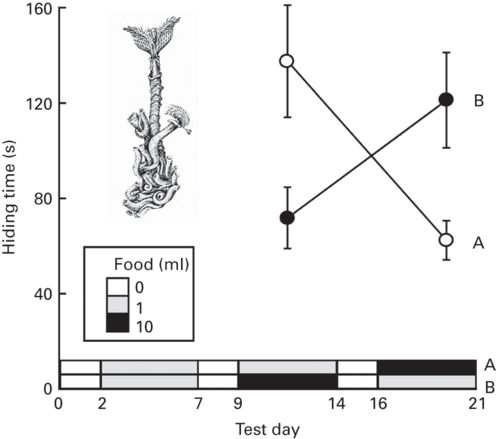
Figure 9.5
Effects of loss of opportunities for foraging by polychaete worms, Serpula vermicularis, on time spent retreated inside their calcareous tubes. Hiding times (mean ± SE) of two groups of serpulids before and after a change in food availability (A) from 1 ml of food suspension to 10 ml or (B) from 10 ml to 1 ml. Values plotted are the means of the average hiding times in several test days for individual tubeworms in the two treatment groups. The low horizontal bars represent the amount of a standard algal suspension provided daily to each tank.
(Redrawn from Dill & Fraser 1997)
Iberian rock lizards, Iberolacerta cyreni, have shorter hiding times when they have detected food before entering refuge than when they have not (Martín et al. 2003a). This was demonstrated by experimentally introducing a mealworm or control stimulus outside the refuge immediately before a simulated attack (“R” and “RNW” treatments vs. “NF” treatment in Figure 9.6). Avoiding the approaching predator is probably the main priority in the absence of food. However, when a lizard hidden in refuge can observe that the prey that it was trying to capture before the predator attacked (“R” treatment), or a new food item provided after the attack (“RNW” treatment) is available outside the refuge, the benefit expected to be gained by emerging from the refuge is presumably higher than the expected cost due to predation risk. Based on this assessment, lizards rapidly emerge. Furthermore, the success of the encounter with food before the attack, and the added possibility of capturing either a new food item, or one that had been available, but not captured when the lizard hid after the predator appeared, differentially affects the magnitude of the costs of refuge use and, consequently, hiding duration. Lizards that have just eaten a prey before the attack (“E” treatment) have longer hiding times than lizards that have not eaten it (“R” treatment) (Figure 9.6). Lizards with a better nutritional state may delay resuming foraging to decrease predation risk. However, when a new prey is offered after the attack (“ENW” and “RNW” treatments), hiding times decrease greatly in both groups independently of the nutritional state (Martín et al. 2003a; Figure 9.6).
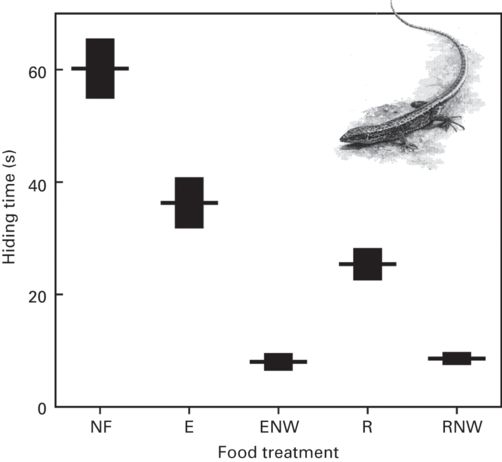
Figure 9.6
Effects of the loss of opportunities for foraging on refuge use by rock lizards, Iberolacerta cyreni. Hiding times (mean ± SE) of lizards after a predatory attack under different experimental treatments of food availability and satiation (NF: no food; E: lizards eat a prey before the attack; ENW: lizards eat a prey before the attack and new food offered after the attack; R: food presented but removed before eating and before the attack; RNW: food removed before eating but new food offered after the attack.
(Redrawn from Martín et al. 2003a)
Similarly, hiding times of striped plateau lizards, Sceloporus virgatus, in rock crevices are shorter when a cricket is placed in front of the crevice opening where it can be seen by the lizard (Cooper 2009a). Food addition also reduces hiding times in fiddler crabs, Uca mjoebergi, but only during periods when food is naturally abundant and foraging is optimal (Reaney 2007). Food additions, however, have no noticeable effect on hiding times of crabs when food levels are naturally low and foraging efficiency is low, probably because the food added only has a weak effect on the benefits of quick emergence after being attacked by a predator. Size of food items near the shell refuge of hermit crabs, Pagurus acadianus, does not affect hiding time (Scarratt & Godin 1992). However, this is consistent with predictions because all food items were larger than could be eaten entirely.
Yellow-bellied marmots emerged sooner from refuges when supplemental food was experimentally placed next to their burrows than when food was absent (Blumstein & Pelletier 2005; Rhoades & Blumstein 2007). However, there is an intriguing interaction; bold individuals that tolerate close approaches before hiding (i.e., with short FIDs) emerge sooner when food is present, while those that are shy and intolerant of approaching humans (i.e., with longer FIDs) take longer to emerge and, in contrast, emerge sooner when food is not present. The latter observation does not apparently fit the expectations of refuge use models. However, it has been suggested (Cooper 2009a) that this finding would be consistent with theory if the placement of food by researchers before the trials in the experimental treatment increased perceived risk more for shy than bold marmots. Another possibility is that individuals that escaped later were hungrier, which would explain why they delayed fleeing and had shorter hiding times. By contrast, those that had longer FIDs might not need the food as much at the moment of attack, but might consider it as a harvestable asset that would reduce foraging costs upon emergence, allowing longer hiding times than when food was absent.
Optimality models predict that prey with greater initial fitness should have longer hiding times (Cooper & Frederick 2007). This is based on the asset protection principle (Clark 1994) that predicts that individuals with greater assets should protect them more carefully. Empirical evidence supports this expectation that prey state (i.e., initial fitness) affects hiding decisions. For example, hungry prey have shorter hiding times. This has been observed in barnacles, Balanus glandula (Dill & Gillett 1991), insects (water striders, Notonecta hoffmani: Sih 1992; N. maculata: Martín & López 2004b), lizards (Iberolacerta cyreni: Martín et al. 2003a), birds (willow tits, Parus montanus: Koivula et al. 1995), and in several species of fish (minnows, Phoxinus phoxinus, and three-spined sticklebacks, Gasterosteus aculeatus: Krause et al. 2000a; Metcalfe & Steele 2001), but not in others (Dowling & Godin 2002). Another observation that supports this prediction is that prey animals with a lower body condition or that have experienced greater weight loss (i.e., that have lower fitness) have shorter hiding times (fish: Krause et al. 1998; lizards: Amo et al. 2007a, b).
Finally, even some plants might adjust for “foraging” cost of being hidden. The plant Mimosa pudica displays a defensive behavior of rapidly folding its leaves when stimulated by touch, thereby decreasing visibility to herbivores, but at the cost of reducing light acquisition and photosynthesis. The time that these plants take to reopen leaves following a disturbance is longer under high light conditions than under more light-limited conditions (Jensen et al. 2011). This suggests that this plant can balance the risk and benefits of antiherbivore behavior in relation to current environmental light conditions.
9.3.2.2 Loss of reproductive opportunities
The loss of reproductive opportunities may also influence hiding times. Many animals are less responsive to predators during the reproductive seasons (Brown & Shine 2004), and this response may differ between sexes. For example, during the mating season, male Iberian rock lizards have shorter hiding times than females at any time and than males outside of the breeding season (Martín et al. 2003b). Furthermore, during the mating season, males have shorter hiding times when a tethered unfamiliar female (i.e., a potential mate) is experimentally placed nearby on their territories territory (Martín et al. 2003b; Figure 9.7). Moreover, in the absence of a female (control situation), males delay emergence when the approach speed of the simulated predator is fast (i.e., high risk) than when it is slow (low risk), but hiding times do not differ between approach speeds or risk levels when a female is present (Figure 9.7). Therefore variation in hiding times seem to reflect different balances between the costs of losing mating opportunities and the benefit of a reduction of predation risk over time.
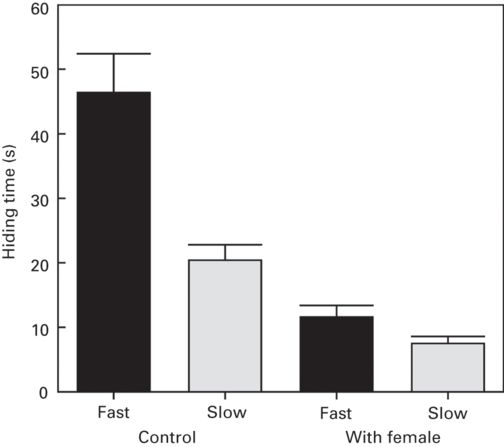
Figure 9.7
Effects of the loss of opportunities for reproduction on refuge use by rock lizards, Iberolacerta cyreni. Hiding times (mean + SE) in a refuge by male lizards in presence or absence (control) of a tethered female after being approached directly by an experimenter at one of two different approach speeds (slow vs. fast) in the mating season.
(Redrawn from Martín et al. 2003b)
Similarly, when mating opportunities for male fiddler crabs, Uca mjoebergi, are experimentally increased by introducing a tethered female, actively courting males engage in highly risky behavior and re-emerge a few seconds after an attack, considerably sooner than males not presented with a female; some males even abandon refuge use completely if a female is present (Reaney 2007). This latter observation confirms the prediction from optimality models (Cooper & Frederick 2007) that prey may risk being killed by leaving the refuge if it may obtain reproductive benefits that are retainable after death.
Hiding time may also be reduced by recent interactions with conspecific competitors due to an increased perceived need to defend feeding or reproductive opportunities, given that a hiding territorial resident is unable to monitor its territory or defend it from conspecific intrusions. The presence of an intruder in the near past can indicate an increased probability of future intrusions. Therefore following a conspecific intrusion, territorial residents should minimize costs from future intrusions at the cost of higher predation risks (Díaz-Uriarte 2001). Thus when a predator attack occurs immediately after an agonistic conspecific interaction, foraging backswimmers, Notonecta maculata, hide underwater, but resume feeding positions at the water surface more quickly and closer to the original position from which they were disturbed, suggesting short-term defense of particular foraging positions (Martín & López 2004b). When male lava lizards, Tropidurus hispidus, have recently encountered a rival male, they emerge sooner after a simulated predator attack, presumably to avoid losing mating opportunities (Díaz-Uriarte 1999, 2001). Similarly, territorial male water skinks, Eulamprus heatwolei, are less likely than non-territorial floaters to hide in refuges and return faster to a basking site after an attack. This may be explained by the trade-off that territorial lizards face between territorial defense and antipredator behavior (Stapley & Keogh 2004). Also, “singing” males of the acoustic pyralid moth, Achroia grisella, within leks respond with shorter pauses in their ultrasonic mating call in response to bat sounds than solitary individuals (Brunel-Pons et al. 2011).
9.3.2.3 Physiological costs
The physiological costs of refuge use on hiding times should be influenced by temperature (in ecotherms), and by oxygen concentration in the water (in aquatic animals). In ectotherms, such as reptiles, the attainment and regulation of an optimal body temperature is essential to maximize physiological and ecological processes, which may conflict with costs expected due to predation risk (Christian & Tracy 1981; Huey 1982). When a lizard or other ectothermic animal hides in a cold refuge (e.g., a rock crevice), its body temperature will quickly fall below optimal values (Polo et al. 2005). This may affect general physiological performance, such as somatic growth (Martín & López 1999b; Amo et al. 2007a,b), and may increase susceptibility to predators after emergence due to a reduced escape speed at low body temperatures (Christian & Tracy 1981). Therefore hiding decisions of ectotherms should reflect the thermal costs of refuge use.
Theoretical models predict, and empirical studies have shown, that as the difference between temperatures inside cold refuges and outside environmental temperatures increases (i.e., when the physiological cost of refuge use increases), hiding times decrease in rock lizards, Iberolacerta cyreni (Martín & López 1999a; Polo et al. 2005; see Figures 9.1 and 9.2), wall lizards, Podarcis muralis (Amo et al. 2003), and striped plateau lizards, Sceloporus virgatus (Cooper & Wilson 2008; Cooper 2009a, 2011a). A lizard’s body temperature that before entering the refuge was correlated with the outside environmental temperature, will decrease more quickly and to a greater extent inside the refuge when the difference in ambient temperatures between the exterior and the refuge is greater. The empirical results suggest that optimization of refuge-use strategies may help lizards to cope with changes in predation risk without incurring excessive physiological costs associated with a lowered body temperature.
Predictions for the effect of difference between body and refuge temperatures in ectotherms are similar in all models, and optimality models predict that hiding times should be strongly affected by the cooling rate inside the refuge (Cooper and Frederick 2007). In lizards, ontogenetic changes in body size affect thermal exchange rates, with smaller individuals having a lower thermal inertia (i.e., they heat and cool faster). This simple physical property may have consequences for thermoregulation, and also for antipredator behavior. Therefore ontogenetic differences in thermal properties might affect costs of refuge use. In comparison with large adults, smaller juvenile rock lizards, Iberolacerta cyreni, delay fleeing to a refuge and have shorter hiding times in cold refuges, probably because their costs of hiding are higher, because they experience faster cooling rates than adults (Martín & López 2003b).
The effect of temperature on hiding decisions may be more complicated in ectothermic animals because they may partially emergence to assess risk prior to complete emergence. In such cases, hiding time has two components. First, appearance time is the time between refuge entry and appearance of a prey without fully emerging. Second, waiting time is the time between appearance and full emergence.
In Schreiber’s green lizards, Lacerta schreiberi, appearance time increases as temperature outside the refuge increases, body mass increases, and temperature inside the refuge decreases (Martín & López 2010). Waiting time is longer when temperature in the refuge is lower, but is not affected by exterior temperature or body mass. Consequently, the total hiding time before complete emergence is longer when the exterior temperature is higher and the interior temperature is lower. The results show that interior and exterior temperatures have opposite effects on appearance time, and that waiting time increases as interior temperature decreases. The increase in appearance time as exterior temperature and body mass increase are consistent with the hypothesis that lizards can spend longer in refuge without suffering great loss of performance capacity when their body temperatures are higher and cooling rates are lower when they enter refuges. Warmer lizards with slower cooling rates take longer to reach a critically low body temperature requiring partial emergence. The increase in appearance time as interior temperature decreases is opposite to previous findings that hiding time increases as interior temperature increases.
Because waiting time also increases as interior temperature decreases, hiding time is longer when refuge temperature is lower and exterior temperature is higher (Martín & López 2010). If lizards allow body temperature to fall lower before appearing at the refuge’s entrance when refuge temperature is lower, cooler lizards may require more time to assess risk prior to full emergence. Furthermore, body temperature might increase during the waiting interval while lizards are partially emerged. This is more likely if the lizards can bask, but also happens because the environmental temperature at the entrance may be higher than the temperature deeper in the refuge. The longer waiting times of lizards that enter cooler refuges may account for the absence of any effect of body mass on hiding time despite the increase in appearance time as body mass increases. Alternatively, lizards having greater thermal inertia take longer to cool, but also take longer to warm, which may lead to countervailing effects on appearance time and waiting time. Although only interior temperature is significantly related to waiting time, possible differences in body temperature upon emerging between smaller and larger lizards could obscure a relationship between body mass and waiting time.
Further research is needed to develop a comprehensive understanding of these findings. However, it is clear that when making decisions regarding refuge use, Lacerta schreiberi lizards seem to consider the physiological costs of being at low temperatures and also the risk of emerging with low escape performance that results from low body temperature.
In some cases, hiding in refuges may preclude breathing or make it difficult to breathe. This too can increase the physiological costs of refuge use. For example, air-breathing dwarf gouramis fish, Colisa lalia, increase their air-breathing frequency at low dissolved oxygen concentrations in water. These gouramis increase their use of refuges (submerged vegetation) and thus decrease their frequency of surfacing events to breathe air in the presence of predatory fish (Wolf & Kramer 1987). The freshwater clam, Corbicula fluminea, protects its soft tissues from small predators by closing its protective valves. This reduces predation risk, but ventilation and oxygen uptake are suspended. Clams reopen their valves sooner when under lower than higher oxygen concentrations; a finding that suggests that hypoxia increases costs of refuge use and increases vulnerability to predation (Saloom & Duncan 2005). Mosquito pupae, Culex pipiens, get aerial oxygen by sticking their air siphons above water. When undisturbed, pupae rest at the surface, but react to physical and visual disturbance by swimming down. The time interval between two consecutive surfacing events (hiding time) is longer under high risk, suggesting, but not demonstrating, a possible trade-off between risk and tolerance of oxygen deficit (Rodríguez-Prieto et al. 2006). Nevertheless, time spent on the surface does not vary with risk levels.
9.3.2.4 Multiple types of predators and conflicting refuge use
A prey’s defenses against one predator may put it at greater risk of being killed by other predators. Some types of refuges may be useful only against particular types of predators or may expose prey to different predators (Sih et al.1998). For example, mortality of mayfly larvae in the presence of both predaceous fish and stonefly larvae is greater than expected because stoneflies in refuges cause mayflies to come out of hiding, thus increase their exposure to fish (Soluk 1993).
Prey exposed to multiple types of predators can experience conflicts. Enhanced survival from one predator may simultaneously increase vulnerability to another predator, especially if prey can deploy defense against only one type of predator at a time (Sih et al. 1998). In such cases, prey should use tactics to defend against the most dangerous predator and ignore the less dangerous predators (McIntosh & Peckarsky 1999; Bouwma & Hazlett 2001). In many cases prey should decide to use a refuge against an exterior predator if the risk of being captured by this predator is higher than the risk of being captured by another type of predator inside the refuge.
Wall lizards, Podarcis muralis, respond to simulated predatory attacks of birds and mammals by hiding inside the nearest rock crevice, but this may expose them to increased risk of predation by the saurophagous smooth snake, Coronella austriaca, which ambushes lizards inside rock crevices. Thus wall lizards employ different alternative escape strategies in relation to their reliance on refuge safety (Amo et al. 2005). Lizards basking close to refuges that they have used recently, hide in them again when attacked. By contrast, lizards that are moving do not always enter the closest refuge, but often run away without hiding. This behavior may be a strategy to avoid entering refuges in which the absence of predators has not been recently ascertained.
Similarly, the gecko Oedura lesueurii uses the same types of refuges as one of its main snake predators, Holocephalus bungaroides. These geckos use their chemosensory ability to avoid entering rock crevices covered with the scent of H. bungaroides (Downes & Shine 1998). Log skinks, Pseudemoia entrecasteauxii, use snake-scented refuges less than predator-free refuges (Stapley 2004). However, when skinks are exposed to risk of predation by a snake inside the refuge and by a bird in the open, skinks do not decrease their use of snake-scented refuges (Stapley 2004), suggesting that predation by birds is more dangerous than the risk of encountering a snake inside a refuge.
Hiding time in refuges should depend on the probability that another type of potential predator is in the refuge. When a refuge is potentially unsafe (e.g., because the prey detects predator cues inside), the probability of being detected by a second type of predator hidden in that refuge increased with time spent in the refuge. Hence, a prey hidden in an unsafe refuge will have increased costs of refuge use and should emerge sooner than from a predator-free refuge. However, the presence of different predator cues may modify a prey’s reliance on refuge safety by altering its assessment of the risk associated with entering a refuge as well as the cost of remaining in one in relation to the risk outside the refuge. For example, after a simulated attack outside a laboratory refuge, wall lizards hid longer when there were no snake cues inside than when chemical or visual snake cues were added to the refuge. Moreover, when visual and chemical snake cues were combined, hiding times were even shorter, supporting an additive effect of several cues to risk assessment (Amo et al. 2004).
9.4 Hiding behavior when in morphological and constructed refuges
Many animals flee to refuges, but are also able to escape by fleeing without hiding, to choose the refuge type, and to change refuges after persistent attacks (Cooper 1998; Martín & López 1999a, 2003a). Other species that are relatively sessile, such as polychaete tubeworms, caddis-fly larvae, mollusks, and barnacles, can only take refuge in a protective structure surrounding their bodies (Dill & Gillett 1991; Johansson & Englund 1995; Dill & Fraser 1997). An intermediate situation occurs when animals have morphological or protective structures that provide partial protection, but are also able to escape actively to safer refuges in their habitat. Examples are hermit crabs, hedgehogs, pangolins, porcupines, armadillos, some fishes, skin-armoured lizards (e.g., the armadillo girdled lizard, Cordylus cataphractus; horned lizards, Phrynosoma spp.) and turtles (McLean & Godin 1989; Doncaster 1993; Losos et al. 2002; Mima et al. 2003; Martín et al. 2005). However, factors that modulate refuge use in these animals remain largely unknown, and characteristics of such prey could complicate escape and hiding decisions predicted by optimal refuge use theory (Martín & López 1999a; Hugie 2003; Cooper & Frederick 2007).
Hermit crabs, Pagurus filholi, have two antipredator tactics: taking refuge in their shells and fleeing. When hermit crabs are dropped into seawater containing chemical stimuli from a predatory crab, they prefer hiding in shells of species that provide more effective protection against crab predators, and hiding times are shorter in the shells of mollusks where they are more vulnerable. However, hermit crabs have shorter hiding times in shells and switch to active fleeing earlier when predator stimuli are present than absent (Mima et al. 2003; Figure 9.8).
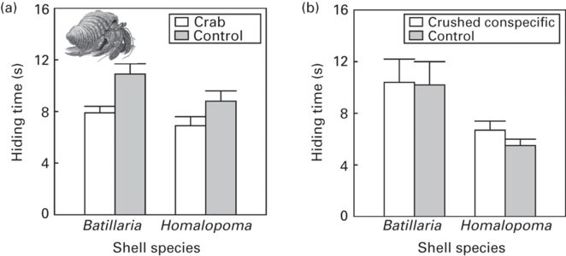
Figure 9.8
Hiding time before active fleeing by hermit crabs, Pagurus filholi, in the shells of Batillaria cumingi (heavy and highly protective) or Homalopo masangarense (light and vulnerable) after being dropped into seawater control or seawater containing (a) stimulus of a predatory crab or (b) stimulus of crushed conspecific.
(Redrawn from Mima et al. 2003)
Spanish terrapins, Mauremys leprosa, adjust their hiding behavior by assessing the risk of emerging before the predator has left the area. Turtles hide longer in their shells when risk level is higher, as indicated by the predator’s behavior (i.e., longer handling duration during the attack, proximity as the turtle hides, and persistence after the attack; Martín et al. 2005). However, because the shell offers only partial protection, increasing hiding time may also increase the risk that the predator is able to injure or kill the turtle. Therefore the possibility of switching from hiding to active escape to a safer refuge (e.g., into water) also shortens hiding times.
In turtles, interspecific differences in hiding behavior appear to be related to the presence of different types of predators in different habitats (Martín et al. 2005; Polo-Cavia et al. 2008). After an attack by a terrestrial predator, basking sliders, Trachemys scripta, remain hidden inside the shell for long periods delaying or avoiding active escape toward the water (Polo-Cavia et al. 2008). In contrast, basking Spanish terrapins, Mauremys leprosa, typically flee toward the safety of deep water immediately, remaining hidden in the shell only when they are far from water (Martín et al. 2005). Sliders hide in the shell before entering water because diving may expose them to dangerous aquatic predators, but for M. leprosa fleeing to water immediately is favored because aquatic predators are relatively uncommon in their habitat (Martín et al. 2005; Polo-Cavia et al. 2008).
9.5 Hiding under simultaneous risks and costs
During an interaction with a predator, prey must simultaneously assess risks and costs for multiple risk factors and several types of costs, and adjust hiding time accordingly. For example, a prey might accurately assess risk level based on predator behavior during the attack, increasing hiding time with increasing directness and approach speeds, but the prey has to simultaneously assess the risk posed by a predator waiting close to the refuge to avoid being captured immediately upon emergence. Conversely, if the prey assesses risk associated with the predator’s distance to the refuge, but fails to consider the predator’s approach speed, it might be captured due to misjudging the predator’s speed.
The “threat-sensitivity hypothesis” (Helfman 1989) predicts that multiple risk factors have additive effects on the intensity of antipredatory responses. This implies that all predator traits contribute to risk in an additive fashion and simultaneous risks must therefore have additive effects (Smith & Belk 2001). However, optimal refuge use theory suggests that interaction between factors should be more common (Cooper & Frederick 2007; Chapter 2): additive effects are predicted to occur only if the differences in curves of risk diminution with time for different risk levels of one risk factor are identical to those for the second risk factor. Effects should be interactive when risk decreases faster with time for one factor than the other, which is a more likely situation given the different characteristics and effects of each risk factor.
A review of empirical studies examining effects of multiple risks and costs on FID and hiding time (Cooper 2009b) revealed far more frequent interactive than additive effects between two risk factors, as predicted by optimality theory. For example, approach speed during the attack and predator proximity after the attack have additive effects on hiding times of rock lizards, but approach speed and directness interact with predator persistence (Martín & López 2004a), with place of emergence (Martín & López 1999a), and with the peculiar characteristics of each individual predator (Cooper et al. 2003). Similarly, in wall lizards, directness interacts with proximity and with persistence of the predator (Martín & López 2005), and in Balearic lizards approach speed interacts with both directness of approach and predator proximity (Cooper et al. 2010) (see Figure 9.3). In Spanish terrapins, risk level (handling duration) and proximity of the predator interacts with the microhabitat where the attack occurs because hiding time inside the shell increases with risk on land, but not in water where emergence could facilitate an alternative active escape (Martín et al. 2005).
However, additive and interactive effects seem to be equally common when a risk and a cost factor are considered simultaneously (Cooper 2009b). In this case, if there are unequal slopes of the risk curves for different risk levels, the cost curves for different magnitudes of costs, or both, we could expect an interaction between risks and costs. If curves for different levels of risks or costs have equal slopes, or differences are too small to be noted, we could expect additive effects. For example, risk due to lower escape ability after tail loss and thermal costs of refuge use have additive effects on hiding time in striped plateau lizards (Cooper & Wilson 2010). Thermal costs interact with directness and speed of the approach in lizards (Martín & López 1999a, 2005), but when there are persistent repeated attacks of a predator, the effects are additive with thermal costs (Polo et al. 2005). More studies of diverse taxa are required to determine the shapes of risk and cost curves. These are needed by optimality models of refuge use to predict joint effects of multiple risks and costs.
9.6 Repeated attacks and multiple hiding decisions
9.6.1 Optimal multiple hiding decisions
In some circumstances, if predators remain waiting outside the refuge and try new attacks or if predator density increases, a particular prey may suffer successive attacks in a short time. Successive attacks may be assessed as indicating increase in the risk of predation, but the costs of refuge use also increase with time spent in the refuge. Thus prey should make new, but related, decisions on when to emerge after each attack. In a field experiment, rock lizards, Iberolacerta cyreni, were approached ten times in a short interval. Hiding times increased progressively with successive attacks, but only when the thermal cost of refuge use was low, as lizards tended to maintain or even to decrease hiding times between approaches when cost of refuge use increased (Martín & López 2001).
Polo et al. (2005) modified previous economic models of hiding time to include the case of a high, sustained level of predation risk during repeated attacks. The model assumes that prey perceive that the probability of a new attack decreases more slowly with time after each successive attack, that costs of refuge use increase with time at the same rate after each attack, and that hiding cost does not start from zero except after the first attack, but accumulates across approaches and begins each time at the maximum level reached in the previous episode of hiding (Figure 9.9). The model predicts that the optimal hiding time should increase following successive attacks and that this increase should not be linear but accelerating (i.e., the rate of increase in hiding time increases over successive emergences).
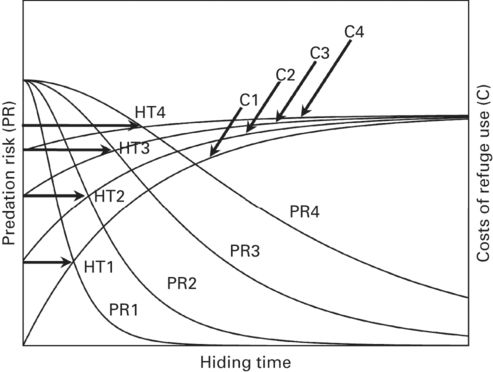
Figure 9.9
An economic model to predict the hiding times of lizards in a refuge after successive continuous attacks as a function of the expected fitness effects of the diminution of the risk of predation in the exterior with time (PR) after each attack (successive PR curves), and variations with time in costs (C) of refuge use after each attack (successive C curves). The optimal hiding times (HT) after each successive attack are shown.
(Redrawn from Polo et al. 2005)
In a laboratory experiment using male Iberian rock lizards, predation risk was identical in two treatments in which temperature inside the refuge was high or low. As predicted, successive hiding times increased in an accelerating rather than linear manner and were shorter when thermal cost of refuge use was higher (Polo et al. 2005). This is similar to previous field observations (Cooper 1998; Martín & López 1999a, 2001). However, in the laboratory, potential confounding variables were held constant and changes from low to high thermal cost were observed consistently in the same individuals.
Assessed risk presumably was higher in the laboratory study (Polo et al. 2005) than in the field (Martín & López 2001) because only one refuge was available in a terrarium and persistent attacks were made from close range in the laboratory, whereas multiple refuges were present and attacks were launched from longer distance in the field. Greater risk explains the increase in laboratory hiding times even when thermal cost was high. Alternatively stated, when predation risk is very high, prey may adopt a conservative strategy and remain inactive in the refuge for very long periods and conserve energy (Polo et al. 2005). But this strategy is dependent on temperature inside the refuge. When refuge temperature is low, lizards should initially emerge as soon as possible to avoid heat loss. However, after repeated attacks, it might be more favorable to remain inactive until risk decreases because emerging with a low body temperature, and thus having poor locomotor performance, may be more dangerous than risking starvation by remaining in the burrow. In contrast, in a refuge with a high temperature, where body temperature does not decrease, prey can resume normal activity immediately after emerging (Polo et al. 2005).
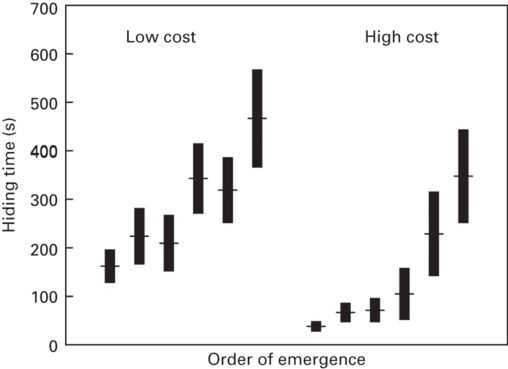
Figure 9.10
Multiple hiding decisions by rock lizards Iberolacerta cyreni. Successive hiding times in a refuge (mean ± SE) after multiple predatory attacks and under low or high thermal costs of refuge use.
(Redrawn from Polo et al. 2005)
9.6.2 Risk assessment affecting multiple hiding decisions
Although not responding to predator cues appropriately can be lethal, an excessive or unnecessary hiding response also can have detrimental effects for prey (Rhoades & Blumstein 2007). Thus when there are multiple persistent predatory attacks it is important to determine whether prey use fixed behaviors or respond flexibly to short-term changes in perceived predation risk by changing their hiding times.
Prey might assess predation risk during successive attacks in several ways. (1) Fixed independent estimation: prey might estimate that each successive attack is independent of the previous one and adjust hiding time to the current risk level in each attack without considering previous risk estimates. (2) Accumulative dependent estimation: prey might consider that each successive attack represents an increase in risk level with respect to the previous estimation regardless of the actual risk level of each single attack. Thus successive hiding times would increase. (3) Flexible dependent estimations: dynamic models have suggested that animals will show tolerance to imperfect information (Bouskila & Blumstein 1992), but that the extent of this tolerance may change from one situation to the next, and that the rules of thumb used to assess risk should be flexible enough to correspond to current conditions (Koops & Abrahams 1998). After a second successive attack indicating predator persistence, it may be necessary to assess risk more accurately. An individual might remember its estimate of the risk level in one attack, compare it with the estimated risk in the current attack, and modify hiding time accordingly. If the second risk level is estimated to be higher than the previous one, prey might consider that it has underestimated the previous risk level and increase hiding time more than expected. If the second risk level is estimated to be lower, then prey might consider that it has overestimated the first risk level and show shorter hiding times than expected. A more conservative option might be to maintain at least the response to the risk level estimated in the first attack, even if the second attack was estimated to be of lower intensity. This could be a viable strategy because overestimation of risk may have milder fitness consequences than underestimating danger (Bouskila & Blumstein 1992; Rhoades & Blumstein 2007; but see Abrams 1994). Alternatively, in some cases, prey may habituate and reduce its antipredator responses to a potential predator that ceases to be regarded as dangerous after repeated non-threatening exposures to it (e.g., Hemmi & Merkle 2009).
In an experimental field study, Martín and López (2004a) examined how short-term changes in risk level of two successive attacks affect successive hiding times of rock lizards (Figure 9.11). The lizards seem to use flexible dependent estimations of risk in each attack. They change the duration of their hiding times through successive attacks as a function of risk level of each attack, but previous estimations of risk are used to assess the new risk level in a second attack. Thus when both attacks are of low intensity, hiding times are of similar short duration after both attacks. When risk increases from the first attack to the second, or both attacks are of high risk, lizards increase hiding times after the second attack. However, when risk level is high initially but decreases for the second attack, lizards maintain the hiding time required for the risk level assessed in the first attack. This might seem to imply overestimation of risk, but the cost of ignoring persistent attack may be higher (Bouskila & Blumstein 1992).
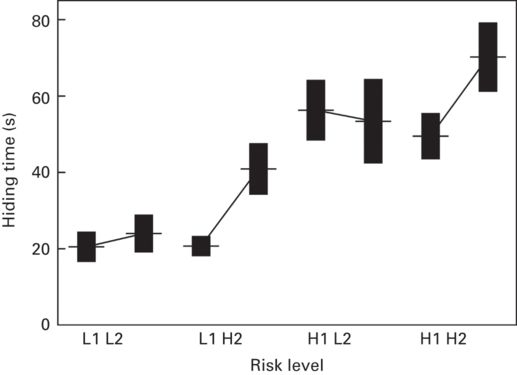
Figure 9.11
Multiple hiding decisions by rock lizards, Iberolacerta cyreni. Successive hiding times in a refuge (mean ± SE) in four treatments in which two successive predatory attacks were both of low risk level (L1L2), increasing in risk level (L1H2), decreasing in risk level (H1L2), or both of high risk level (H1H2).
(Redrawn from Martín & López 2004a)
In contrast, wall lizards, Podarcis muralis, seem to consider each of three successive attacks to be independent. They adjust hiding time to current risk in each attack without considering the risk levels in previous attacks (Martín & López 2005). This indicates that lizards may track short-term changes in risk level through time and modify their initial responses when required. This might occur if lizards considered that a predator should usually not outwait the prey after an unsuccessful attack (Hugie 2003), but depart to look for other potential prey, which is plausible because lizard density is very high in this population. For this species, two successive attacks are likely to come from different individual predators, in which case risk may be assessed again in each attack independently of previous estimates.
9.6.3 Monitoring from the refuge to determine hiding time
Most studies of refuge use have analyzed situations in which information on predator behavior is unavailable to hiding prey. In this case current risk of being killed upon emergence can only be estimated by prey based on time elapsed since the attack and characteristics of the attack (Sih 1992; Martín & López 1999a; Hugie 2003). In other cases, prey can monitor predators from the refuge safely and use information gained to decide hiding times (Cooper 2008; Polo et al. 2011). Nevertheless, monitoring predators from the refuge entails costs, such as revealing the refuge’s position to the predator while scanning for the predator presence.
When monitoring behavior is possible for lizards at, or partially outside, the refuge’s entry, hiding time, as noted previously, can be divided into appearance time and waiting or monitoring time. These different times may be differentially affected by risk and costs of refugia.
For example, in response to series of simulated attacks (low vs. high risks), rock lizards, Iberoalcerta cyreni, have appearance times that do not differ between risk levels, but have longer monitoring times after a single low-risk approach of the predator than after a direct unsuccessful attack (Polo et al. 2011). This may occur because, if there has not been a clear attack, uncertainty about future risk would be greater, and prey may need more time before leaving the refuge to ensure that a predator has not detected the lizard and that it is not lying in wait nearby. These results suggest that different levels of uncertainty about future risks, rather than just the previous risk level during the attack, seem to modulate monitoring time from the refuge and, thereby, total hiding times.
9.6.4 Long-term temporal patterns of risk affecting hiding decisions
Prey in nature experience a broad range of temporal patterns of predation risk, and it is not entirely clear how prey respond to these variations (Lima & Bednekoff 1999; Sih et al. 2000). This is important because it has been predicted that antipredator behavior in situations with different levels of predation risk should vary with not just the immediate level of risk but also with the preceding temporal pattern of perceived risk (i.e., risk allocation hypothesis; sensu Lima & Bednekoff 1999).
Rock lizards, Iberolacerta cyreni, were subjected to a series of simulated repeated predatory attacks in the laboratory (24 attacks in a four-hour period) of different risk level (i.e., low vs. high), with similar overall numbers of low-risk and high-risk attacks across a trial, but with three different temporal patterns of presentation (i.e., a series of low-risk approaches and then a series of high-risk attacks, or the opposite pattern, or successive attacks where risk level changed randomly; Martín et al. 2009). Under predictable temporal patterns of risk (i.e., multiple attacks with the same risk level in the recent past) lizards show accurate antipredator responses to each of the successive attacks. However, when risk is random, individuals are not able to predict the level of the next predatory attack and adopt a conservative strategy with longer hiding times than required by the actual current risk level.
On the other hand, hiding times may be shaped by natural selection or maternal effects. These may act through long-term differences in exposure to predation between populations. For example, the first-generation of male crickets, Gryllus integer, reared in the laboratory from field-caught mothers from a high-predation habitat hide longer when placed in a novel environment in the laboratory than male crickets born from mothers from a low-predation habitat (Hedrick & Kortet 2006).Therefore differences in behavior are due to either non-genetic environmental-maternal effects or genetic effects. Similarly, Balearic lizards, Podarcis lilfordi, from two islet populations with different predation pressure differ in hiding time (almost nine times longer under high predation pressure) (Cooper et al. 2009). And wild-caught guppies, Poecilia reticulata, from high-predation localities emerge sooner from shelter compared to those from low-predation localities (Harris et al. 2010).
9.7 Conclusions and future directions
Many predictions from theoretical models of hiding time have been successfully tested using simply designed studies that measured hiding behavior, but many topics require research in the future. First, the emphasis of available studies is heavily on lizards. Studies of a wide range of different taxa, in different environments and with different physiological and morphological constrains may impart more useful knowledge and help to assess the range of applicability of the predictions of theoretical models. Second, new, probably more elaborated and complex, models and experiments are needed to understand hiding behavior in situations when several risk and costs factors act simultaneously and when mutliple responses depend of previous decisions. In addition, variation in decisions about refuge use may interact with other antipredatory behaviors, such as FID and DF, and may also depend on differences in personality that are not considered in current models. Encounters with predators often involve multiple, complex successive chases and escape sequences. In such cases, prey should continously assess and track changes in risk level, consider potential accumulative costs of refuge use, and estimate the relative success of its previous antipredatory decisions to modify them accordingly. The ability to monitor predators and quality of the information about predator behavior and “intentions” have been considered little. Although the waiting-game model (Hugie 2003) predicts that hiding time is longer than a predator’s giving-up time, very little empirical attention has been paid to understanding why predators should not be able to predict hiding times of their prey better (as human researchers seem to do) and counteract their antipredatory strategies. This points out the need for models and studies that consider the actions of both prey and predator behavior simultaneously. New models and experiments using hiding time and related variables will lead to a greater understanding of many aspects of risk-taking decisions.
References
Abrams, P. A. (1994). Should prey overestimate the risk of predation? American Naturalist, 144, 317-328.
Amo, L., López, P. & Martín, J. (2003). Risk level and thermal costs affect the choice of escape strategy and refuge use in the wall lizard, Podarcis muralis. Copeia, 2003, 899-905.
Amo, L., López, P. & Martín, J. (2004). Wall lizards combine chemical and visual cues of ambush snake predators to avoid overestimating risk inside refuges. Animal Behaviour, 67, 647-653.
Amo, L., López, P. & Martín, J. (2005). Flexibility in antipredatory behavior allows wall lizards to cope with multiple types of predators. Annales Zoologici Fennici, 42, 109-121.
Amo, L., López, P. and Martín, J.(2007a). Refuge use: a conflict between avoiding predation and losing mass in lizards. Physiology and Behavior, 90, 334-343.
Amo, L., López, P. & Martín, J. (2007b). Pregnant female lizards Iberolacerta cyreni adjust refuge use to decrease thermal costs for their body condition and cell mediated immune response. Journal of Experimental Zoology A, 307, 106-112.
Bateman, P. W. & Fleming, P. A. (2006). Sex, intimidation and severed limbs: The effect of simulated predator attack and limb autotomy on calling and emergence behaviour in the field cricket Gryllus bimaculatus. Behavioral Ecology and Sociobiology, 59, 674-681.
Blumstein, D. T. & Pelletier, D. (2005). Yellow-bellied marmot hiding time is sensitive to variation in costs. Canadian Journal of Zoology, 83, 363-367.
Bouskila, A. & Blumstein, D. T. (1992). Rules of thumb for predation hazard assessment: Predictions from a dynamic model. American Naturalist, 139, 161-176.
Bouwma, P. & Hazlett, B. A. (2001). Integration of multiple predator cues by the crayfish Orconectes propinquus. Animal Behaviour, 61, 771-776.
Brown, G. P. & Shine, R. (2004). Effects of reproduction on the antipredator tactics of snakes (Tropidonophis mairii, Colubridae). Behavioral Ecology and Sociobiology, 56, 257-262.
Brunel-Pons, O., Alem, S. & Greenfield, M. D. (2011). The complex auditory scene at leks: Balancing antipredator behaviour and competitive signalling in an acoustic moth. Animal Behaviour, 81, 231-239.
Cabido, C., Galán, P., López, P. & Martín, J. (2009). Conspicuousness-dependent antipredatory behavior may counteract coloration differences in Iberian rock lizards. Behavioral Ecology, 20, 362-370.
Christian, K. A. & Tracy, C. R. (1981). The effect of the thermal environment on the ability of Galapagos land iguanas to avoid predation during dispersal. Oecologia, 49, 218-223.
Clark, C. W. (1994). Antipredator behavior and the asset protection principle. Behavioral Ecology, 5, 159-170.
Cooper, W. E., Jr. (1998). Risk factors and emergence from refuge in the lizard Eumeces laticeps. Behaviour, 135, 1065-1076.
Cooper, W. E., Jr. (2008). Visual monitoring of predators: occurrence, cost and benefit for escape. Animal Behaviour, 76, 1365-1372.
Cooper, W. E., Jr. (2009a). Theory successfully predicts hiding time: new data for the lizard Sceloporus virgatus and a review. Behavioral Ecology, 20, 585-592.
Cooper, W. E., Jr. (2009b). Fleeing and hiding under simultaneous risks and costs. Behavioral Ecology, 20, 665-671.
Cooper, W. E., Jr. (2009c). Variation in escape behavior among individuals of the striped plateau lizard Sceloporus virgatus may reflect differences in boldness. Journal of Herpetology, 43, 495-502.
Cooper, W. E., Jr. (2011a). Risk, escape from ambush, and hiding time in the lizard Sceloporus virgatus. Herpetologica, 68, 505-513.
Cooper, W. E., Jr. (2011b). Escape strategy and vocalization during escape by American Bullfrogs (Lithobates catesbeianus). Amphibia-Reptilia, 32, 213-221.
Cooper, W. E., Jr. & Avalos, A. (2010). Predation risk, escape and refuge use by mountain spiny lizards (Sceloporus jarrovii). Amphibia-Reptilia, 31, 363-373.
Cooper, W. E., Jr. & Blumstein, D. T. (2014). Novel effects of monitoring predators on costs of fleeing and not fleeing explain flushing early in economic escape theory. Behavioral Ecology, 25, 44-52.
Cooper, W. E., Jr. & Frederick, W. G. (2007). Optimal time to emerge from refuge. Biological Journal of The Linnean Society, 91, 375-382.
Cooper, W. E., Jr. & Wilson, D. S. (2008). Thermal cost of refuge use affects refuge entry and hiding time by striped plateau lizards Sceloporus virgatus. Herpetologica, 64, 406-412.
Cooper, W. E., Jr. & Wilson, D. S. (2010). Longer hiding time in refuge implies greater assessed risk after capture and autotomy in striped plateau lizards (Sceloporus virgatus). Herpetologica, 66, 425-431.
Cooper, W. E., Jr., Martín, J. & López, P. (2003). Simultaneous risks and differences among individual predators affect refuge use by a lizard, Lacerta monticola. Behaviour, 140, 27-41.
Cooper, W. E., Jr., Hawlena, D. & Pérez-Mellado, V. (2009). Islet tameness: Escape behavior and refuge use in populations of the Balearic lizard (Podarcis lilfordi) exposed to differing predation pressure. Canadian Journal of Zoology, 87, 912-919.
Cooper, W. E., Jr., Hawlena, D. & Pérez-Mellado, V. (2010). Influence of risk on hiding time by Balearic lizards (Podarcis lilfordi): Predator approach speed, directness, persistence, and proximity. Herpetologica, 66, 131-141.
Díaz-Uriarte, R. (1999). Anti-predator behaviour changes following an aggressive encounter in the lizard Tropidurus hispidus. Proceedings of the Royal Society of London Series B, Biological Sciences, 266, 2457-2464.
Díaz-Uriarte, R. (2001). Territorial intrusion risk and antipredator behaviour: A mathematical model. Proceedings of the Royal Society of London Series B, Biological Sciences, 268, 1165-1173.
Dill, L. M. & Fraser, A. H. G. (1997). The worm re-turns: hiding behavior of a tube-dwelling marine polychaete, Serpula vermicularis. Behavioral Ecology, 8, 186-193.
Dill, L. M. & Gillet, J. F. (1991). The economic logic of the barnacle Balanus glandula (Darwin) hiding behaviour. Journal of Experimental Marine Biology and Ecology, 153, 115-127.
Doncaster, C. P. (1993). Influence of predation threat on foraging pattern: The hedgehog’s gambit. Revue d Ecologie-la Terre et la Vie, 48, 207-213.
Dowling, L. M. & Godin, J.-G. J. (2002). Refuge use in a killifish: influence of body size and nutritional state. Canadian Journal of Zoology, 80, 782-788
Downes, S. & Shine, R. (1998). Sedentary snakes and gullible geckos: Predator-prey coevolution in nocturnal rock dwelling reptiles. Animal Behaviour, 55, 1373-1385.
Godin, J. G. L. & Sproul, C. D. (1988). Risk taking in parasitized sticklebacks under threat of predation: Effects of energetic need and food availability. Canadian Journal of Zoology, 66, 2360-2367.
Guliam, J. F. & Fraser, D. F. (1987). Habitat selection under predation hazard: Test of a model with foraging minnows. Ecology, 68, 1856-1862.
Harris, S., Ramnarine, I. W., Smith, H. G. & Pettersson, L. B. (2010). Picking personalities apart: estimating the influence of predation, sex and body size on boldness in the guppy Poecilia reticulata. Oikos, 119, 1711-1718.
Hedrick, A. V. (2000). Crickets with extravagant mating songs compensate for predation risk with extra caution. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267, 671-675.
Hedrick, A. V. & Kortet, R. (2006). Hiding behaviour in two cricket populations that differ in predation pressure. Animal Behaviour, 72, 1111-1118.
Helfman, G. S. (1989). Threat-sensitive predator avoidance in damselfish-trumpetfish interactions. Behavioral Ecology and Sociobiology, 24, 47-58.
Hemmi, J. & Merkle, T. (2009). High stimulus specificity characterizes anti-predator habituation under natural conditions. Proceedings of the Royal Society of London Series B, Biological Sciences, 276, 4381-4388.
Huey, R. B. (1982). Temperature, physiology and the ecology of reptiles. In Gans, C. & Pough, F. H. (ed.), Biology of the Reptilia, Vol. 12, pp. 25-91. New York: Academic Press.
Hugie, D. M. (2003). The waiting game: A ‘‘battle of waits’’ between predator and prey. Behavioral Ecology, 14, 807-817.
Jennions, M. D., Backwell, P. R. Y., Mourai, M. & Christy, J. H. (2003). Hiding behaviour in fiddler crabs: How long should prey hide in response to a potential predator? Animal Behaviour, 66, 251-257.
Jensen, E. L., Dill, L. M. & Cahill, J. F., Jr. (2011). Applying behavioral-ecological theory to plant defense: Light-dependent movement in Mimosa pudica suggest a trade-off between predation risk and energetic reward. American Naturalist, 177, 377-381.
Johansson, A. & Englund, G. (1995). A predator-prey game between bullheads and case-making caddis larvae. Animal Behaviour, 50, 785-792.
Journey, L., Drury, J. P., Haymer, M., Rose, K. & Blumstein, D. T. (2013). Vivid birds respond more to acoustic signals of predators. Behavioral Ecology and Sociobiology, 67, 1285-1293.
Koivula, K., Rytkönen, S. & Orell, M. (1995). Hunger dependency of hiding behaviour after a predator attack in dominant and subordinate willow tits. Ardea, 83, 397-404.
Koops, M. A. & Abrahams, M. V. (1998). Life history and the fitness consequences of imperfect information. Evolutionary Ecology, 12, 601-613.
Krause, J., Loader, S. P., McDermott, J. & Ruxton, G. D. (1998). Refuge use by fish as a function of body length-related metabolic expenditure and predation risks. Proceedings of the Royal Society of London. Series B: Biological Sciences, 265, 2373-2379.
Krause, J., Cheng, D. J.-S., Kirkman, E. & Ruxton, G. D. (2000a). Species-specific patterns of refuge use in fish: The role of metabolic expenditure and body length. Behaviour, 137, 1113-1127.
Krause, J., Longworth, P. & Ruxton, G. D. (2000b). Refuge use in sticklebacks as a function of body length and group size. Journal of Fish Biology, 56, 1023-1027.
Lewkiewicz, D. A. & Zuk, M. (2004). Latency to resume calling after disturbance in the field cricket, Teleogryllus ocanicus, corresponds to population-level differences in parasitism risk. Behavioral Ecology and Sociobiology, 55, 569-573.
Lima, S. L. & Bednekoff, P. A. (1999). Temporal variation in danger drives antipredator American Naturalist, 153, 649-659.
Lima, S. L. & Dill, L. M. (1990). Behavioral decisions made under the risk of predation: A review and prospectus. Canadian Journal of Zoology, 68, 619-640.
López, P., Hawlena, D., Polo, V., Amo, L. & Martín, J. (2005). Sources of interindividual shy-bold variations in antipredatory behaviour of male Iberian rock-lizards. Animal Behaviour, 69, 1-9.
Losos, J. B., Mouton, P. le F. N., Bickel, R., Cornelius, I. & Ruddock, L. (2002). The effect of body armature on escape behaviour in cordylid lizards. Animal Behaviour, 64, 313-321.
Martín, J. & López, P. (1999a). When to come out from a refuge: risk-sensitive and state-dependent decisions in an alpine lizard. Behavioral Ecology, 10, 487-492.
Martín, J. & López, P. (1999b). An experimental test of the costs of antipredatory refuge use in the wall lizard, Podarcis muralis. Oikos, 84, 499-505.
Martín, J. & López, P. (2001). Repeated predatory attacks and multiple decisions to come out from a refuge in an alpine lizard. Behavioral Ecology, 12, 386-389.
Martín, J. & López, P. (2003a). Changes in the escape responses of the lizard Acanthodactylus erythrurus under persistent predatory attacks. Copeia, 2003, 408-413.
Martín, J. & López, P. (2003b). Ontogenetic variation in antipredatory behavior of Iberian-rock lizards (Lacerta monticola): Effects of body-size-dependent thermal-exchange rates and costs of refuge use. Canadian Journal of Zoology, 81, 1131-1137.
Martín, J. & López, P. (2004a). Iberian rock lizards (Lacerta monticola) assess short-term changes in predation risk level when deciding refuge use. Journal of Comparative Psychology, 118, 280-286.
Martín, J. & López, P. (2004b). Balancing predation risk, social interference and foraging opportunities in backswimmers, Notonecta maculata. Acta Ethologica, 6, 59-63.
Martín, J. & López, P. (2005). Wall lizards modulate refuge use through continuous assessment of predation risk level. Ethology, 111, 207-219.
Martín, J. & López, P. (2010). Thermal constraints of refuge use by Schreiber’s green lizards, Lacerta schreiberi. Behaviour, 147, 275-284.
Martín, J., López, P. & Cooper, W. E., Jr. (2003a). When to come out from a refuge: Balancing predation risk and foraging opportunities in an alpine lizard. Ethology, 109, 77-87.
Martín, J., López, P. & Cooper, W. E., Jr. (2003b). Loss of mating opportunities influences refuge use in the Iberian rock lizard, Lacerta monticola. Behavioral Ecology and Sociobiology, 54, 505-510.
Martín, J., Marcos, I. & López, P. (2005). When to come out from your own shell: Risk sensitive hiding decisions in terrapins. Behavioral Ecology and Sociobiology, 57, 405-411.
Martín, J., López, P. & Polo, V. (2009). Temporal patterns of predation risk affect antipredatory behaviour allocation by Iberian rock-lizards. Animal Behaviour, 77, 1261-1266.
McIntosh, A. R. & Peckarsky, B. L. (1999). Criteria determining behavioural responses to multiple predators by a stream mayfly. Oikos, 85, 554-564.
McLean, E. B. & Godin, J. G. J. (1989). Distance to cover and fleeing from predators in fish with different amounts of defensive armour. Oikos, 55, 281-290.
Metcalfe, N. B. & Steele, G. I. (2001). Changing nutritional status causes a shift in the balance of nocturnal to diurnal activity in European minnows. Functional Ecology, 15, 304-309.
Mima, A., Wada, S. & Goshima, S. (2003). Antipredador defence of the hermit crab Pagurus filholi introduced by predator crabs. Oikos, 102, 104-110.
Polo, V., López, P. & Martín, J. (2005). Balancing the thermal costs and benefits of refuge use to cope with persistent attacks from predators: A model and an experiment with an alpine lizard. Evolutionary Ecology Research, 7, 23-35.
Polo, V., López, P. & Martín, J. (2011). Uncertainty about future predation risk modulates monitoring behaviour from refuges in lizards. Behavioral Ecology, 22, 218-223.
Polo-Cavia, N., López, P. & Martín, J. (2008). Interspecific differences in responses to predation risk may confer competitive advantages to invasive freshwater turtle species. Ethology, 114, 115-123.
Reaney, L. T. (2007). Foraging and mating opportunities influence refuge use in the fiddler crab, Uca mjoebergi. Animal Behaviour, 73, 711-716.
Rhoades, E. & Blumstein, D. T. (2007). Predicted fitness consequences of threat-sensitive hiding behavior. Behavioral Ecology, 18, 937-943.
Rodríguez-Prieto, I., Fernández-Juricic, E. & Martín, J. (2006). Anti-predator behavioral responses of mosquito pupae to aerial predation risk. Journal of Insect Behavior, 19, 373-381.
Saloom, M. E. & Duncan, R. S. (2005). Low dissolved oxygen levels reduce anti-predation behaviours of the freshwater clam Corbicula fluminea. Freshwater Biology, 50, 1233-1238.
Scarratt, A. M. & Godin, J.-G. J. (1992). Foraging and antipredator decisions in the hermit crab Pagurus acadianus (Benedict). Journal of Experimental Marine Biology and Ecology, 156, 225-238.
Sih, A. (1992). Prey uncertainty and the balancing of antipredator and feeding needs. American Naturalist, 139, 1052-1069.
Sih, A. (1997). To hide or not to hide? Refuge use in a fluctuating environment. Trends in Ecology and Evolution, 12, 375-376.
Sih, A., Krupa, J. & Travers, S. (1990). An experimental study on the effects of predation risk and feeding regime on the mating behavior of the water strider, Gerris remigis. American Naturalist, 135, 84-290.
Sih, A., Kats, L. B. & Moore, R. D. (1992). Effects of predatory sunfish on the density, drift and refuge use of stream salamander larvae. Ecology, 73, 1418-1430.
Sih, A., Englund, G. & Wooster, D. (1998). Emergent impacts of multiple predators on prey. Trends in Ecology and Evolution, 13, 350-355.
Sih, A., Ziemba, R. & Harding, K. C. (2000). New insights on how temporal variation in predation risk shapes prey behavior. Trends in Ecology and Evolution, 15, 3-4.
Smith, M. E. & Belk, M. C. (2001). Risk assessment in western mosquito fish (Gambusia affinis): Do multiple cues have additive effects. Behavioral Ecology and Sociobiology, 51, 101-107.
Soluk, D. A. (1993). Multiple predator effects: Predicting combined functional response of stream fish and invertebrate predators. Ecology, 74, 219-225.
Stankowich, T. & Blumstein, D. T (2005). Fear in animals: A review and metaanalysis of risk assessment. Proceedings of the Royal Society of London. Series B: Biological Sciences, 272, 2627-2634.
Stapley, J. (2004). Do log skinks (Pseudemoia entrecasteauxii) modify their behaviour in the presence of two predators? Behavioral Ecology and Sociobiology, 56, 185-189.
Stapley, J. & Keogh, J. S. (2004). Exploratory and antipredator behaviours differ between territorial and nonterritorial male lizards. Animal Behaviour, 68, 841-846.
Werner, E. E., Gilliam, J. F., Hall, D. J. & Mittelbach, G. G. (1983). An experimental test of the effects of predation risk on habitat use. Ecology, 64, 1540-1548.
Wilson, C. D., Arnott, G. & Elwood, R. W. (2012). Freshwater pearl mussels show plasticity of responses to different predation risks but also show consistent individual differences in responsiveness. Behavioural Processes, 89, 299-303.
Wolf, N. G. & Kramer, D. L. (1987). Use of cover and the need to breathe: The effects of hypoxia on vulnerability of dwarf gouramis to predatory snakeheads. Oecologia, 73:127-132.