ESCAPING FROM PREDATORS An Integrative View of Escape Decisions (2015)
Part III Related behaviors and other factors influencing escape
15 The personality of escape
Pilar López and José Martín
15.1 Introduction
Experiments quantifying escape responses have revealed considerable inter-individual variation (Krause & Godin 1996; Jones et al. 2009), which might result from inter-individual differences in the ability to detect predators (“perceptual limit hypothesis”; Quinn & Cresswell 2005a), or in the time devoted to antipredator vigilance (Cresswell et al. 2003). Such inter-individual variation may be adaptive and represent different behavioral strategies (reviewed in Dall et al. 2004; Bell 2007; Réale et al. 2007; Smith & Blumstein 2008; Dingemanse & Wolf 2010). In behavioral and evolutionary ecology, the suites of behavioral traits with consistent individual differences across situations and contexts, or functional behavioral categories (e.g., foraging, social interactions, avoiding predators, etc.), are known as “behavioral syndromes” (Sih et al. 2004a,b; Sih & Bell 2008) or “animal personalities” (Dall et al. 2004; Groothuis & Carere 2005; Carere & Maestipieri 2013). In the past they were also referred to as temperament, coping styles, or coping strategies (Boissy 1995; Wechsler 1995; Koolhaas et al. 1999). Animal personalities have underlying genetic and physiological mechanisms (Koolhaas et al. 1999; Van Oers et al. 2004a), may be heritable (Van Oers et al. 2004b; Bell 2009) and have fitness consequences (Smith & Blumstein 2008).
We review individual differences in escape and hiding responses, and ask how the evolution of different personalities (i.e., the coexistence of different behavioral types in a population) is compatible with the theoretical prediction of a single optimal escape or hiding behavior response. Most work on personality in animals has focused on laboratory and field studies, considering variation in behaviors such as exploration, neophobia, and risk taking (reviewed in Sih et al. 2004a,b; Bell 2007; Réale et al. 2007). Particularly, escape responses and hiding behavior have often been used to characterize the personality of several animals (e.g., Sih et al. 2003; López et al. 2005; Hedrick & Kortet 2006, 2012; Cooper 2009; Carter et al. 2012a,b). Individuals that escape earlier from predators or spend longer times hidden in refuges can be considered as “shy” individuals. By contrast, “bold” individuals delay escape for longer or have shorter hiding times. However, the concept of personality or a behavioral syndrome does not imply a simple dichotomy of behavioral types (e.g., proactive vs. reactive or shy vs. bold), but very often there is a continuous distributionof behavioral types and we should view personalities as existing along a continuum.
15.2 Is escape behavior a personality trait?
Behavioral syndromes or personalities were described as suites of correlated behaviors, with individual consistent differences being expressed in different behaviors in different situations and contexts (Sih et al. 2004a,b). A formal statistical definition, which has recently been proposed, suggests that mixed-effect statistical models can be applied to estimate a suite of between- and within-individual variance components of behavior (Dingemanse & Dochtermann 2013; see also Garamszegi & Herczeg 2012). Thus personalities could be identified if there is consistent repeatability in the inter-individual differences between individuals. In other words, if the degree to which a behavioral trait differs between individuals (i.e., between individual variance) is greater than the degree to which a single observation differs from an individual’s mean (i.e., within-individual variance). Repeatability represents the phenotypic variation attributable to differences between individuals, which is of key importance because it provides a standardized estimate of individuality that can be compared across studies. However, individuals can be consistent in their responses but also be plastic and change their responses between situations following behavioral reaction norms (Nusey et al. 2007; Biro & Stamps 2008; Dingemanse et al. 2010; see Figure 15.1). In that case, mixed-effect models allow the statistical definition of personality and plasticity as behavioral reaction norm intercepts and slopes respectively. When intercepts of these reaction norms differ between individuals in different situations, but slopes did not differ between individuals, we still could talk of personalities maintained across different situations. In addition, we can statistically define behavioral syndromes as non-zero between-individual correlations across behavioral attributes (Dingemanse & Dochtermann 2013).
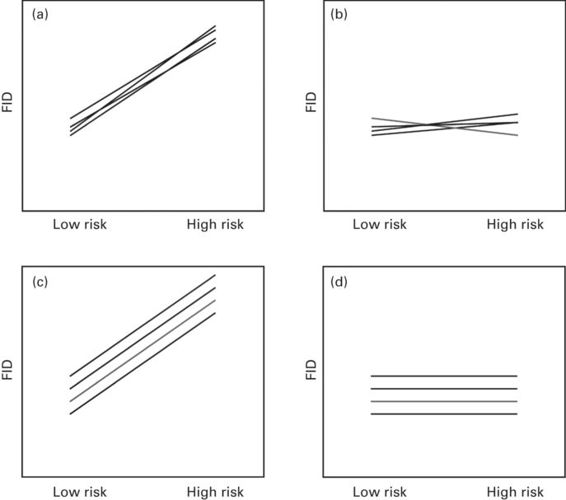
Figure 15.1
Consistency and plasticity in the escape response. Four hypothetical reaction norms of flight initiation distance (FID) responses of several individuals under two risk level situations. (a) The average FID increases when risk is high (plasticity) but there are not consistent personality differences between individuals. (b) The average FID response does not change between risk levels (no plasticity), and there is no consistency in the responses of the same individuals between risk levels. (c) The FID increases similarly in all individuals (plasticity), and the rank order differences between individuals (personality) is maintained consistently in both risk levels. (d) There are no changes in FID between risk levels (no plasticity) but there are consistent inter-individual differences in personality across risk levels.
(Adapted from Nussey et al. 2007)
Consider flight initiation distance (FID). We can imagine four different scenarios (Figure 15.1). On the one hand, individuals may react in a similar way to different risk levels and the average FID of the population may not change between risk levels (i.e., no plasticity; Figure 15.1b,d), but we will have different personalities if the variance of the FID responses of the same individual is consistently lower than the variance in FID between individuals (i.e., individuals differ in intercepts but not in slopes of the reaction norm; Figure 15.1d). We will not find different personalities if there are not consistent inter-individual differences in FID (Figure 15.1b). On the other hand, individuals may be sensitive and plastic and change their escape responses according to risk levels (Figure 15.1a,c), but they will still show different personalities in FID if the rank order differences between individuals is maintained consistently in both risk levels (i.e., individuals differ in intercepts but not in slopes; Figure 15.1c). Finally, individuals may be plastic and change their FID between risk levels, but with a lack of consistent personality differences between individuals (i.e., individuals differ in slopes; Figure 15.1a).
Therefore if escape and hiding behavior were personality traits, we should expect that individuals differed consistently in their responses. However, individuals may show plasticity in different situations (e.g., different risk levels) while maintaining their personality rank in their responses. Moreover, when escaping from predators, bolder individuals might be also expected to show a behavioral syndrome and be bolder in other contexts, such as in conspecific agonistic interactions, foraging, etc. Although these predictions, and the existence of shy-bold escape syndromes, have been rarely tested specifically, we will review the evidence from several personality studies that have used escape and hiding behavior, or related antipredatory behaviors.
15.2.1 Consistency in the escape and hiding responses
To demonstrate individual repeatability, one must study the same subject repeatedly. However, relatively few studies have tested for repeatability of escape behavior in the same individuals in different situations. This is because most studies of escape behavior use independent observations (i.e., different individuals) in different treatments. Also, in most cases, measuring escape behavior requires simulating predatory attacks toward the tested animal. Thus most studies used different individuals to avoid the problem of habituation (see below) or sensitization, where animals decrease or increase, respectively, the magnitude of their subsequent responses to repeated risk situations. However, it would be possible to use a reaction norm approach (Biro & Stamps 2008; Dingemanse & Dochtermann 2013), whereby, by testing individuals repeatedly, it is possible to study both the slope and the intercept of their escape responses and determine whether there are consistent personality differences in responses across situations (see above, Figure 15.1).
One study examined whether reliable individual differences in boldness occur in several aspects of escape and hiding behavior in the striped plateau lizard, Sceloporus virgatus (Cooper 2009). Repeated measures of FID were positively correlated in several tests, indicating that individual differences in boldness were consistent in different situations and over intervals of a few minutes to a day or longer. A similar result was observed for distance fled. Hiding times of the same individuals (two repeated observations in different situations) were correlated in some tests where risk level changed (approach speed, directness, and predator proximity), but not in other tests where risk (successive approaches) or costs levels changed (presence of food outside the refuge; Cooper 2009, 2011). In this case, the latter result could be explained by uncontrolled differences between trials or small effect sizes.
Namibian rock agamas, Agama planiceps, have FIDs that are highly consistent within individuals (between 9 and 15 repeated measures) over a period of almost 50 days, and across situations (seasons). Also, there is more variation in FID between than within individuals, independently of other covariates such as body mass, temperature, and time of day (Carter et al. 2010, 2012a,b; Figure 15.2).
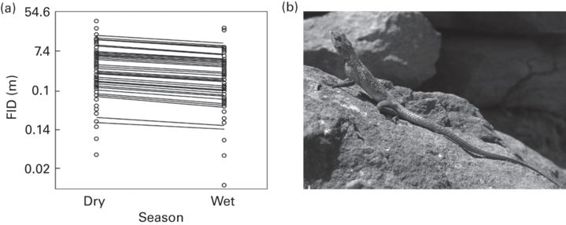
Figure 15.2
Consistency between seasons in the flight initiation distance (FID) of Namibian rock agamas, Agama planiceps. Each point represents an observation of an individual that was observed on multiple occasions in each season. Each line represents each individual’s best linear unbiased predictor for the intercept.
(Figure redrawn from Carter et al. 2012b; photograph by J. Martín)
Similarly, within-individual repeatability in FID is very high in burrowing owls, Athene cunicularia, after controlling for several confounding effects (sex, territory). Furthermore, most individuals do not change their escape responses, and there are as many birds decreasing as increasing their FIDs across successive trials (Carrete & Tella 2010; Figure 15.3). In contrast, in yellow-bellied marmots, Marmota flaviventris, individual identity is not a significant predictor of FID (Runyan & Blumstein 2004). This negative result is probably explained by the very different situational contexts in which individuals were observed in different occasions.
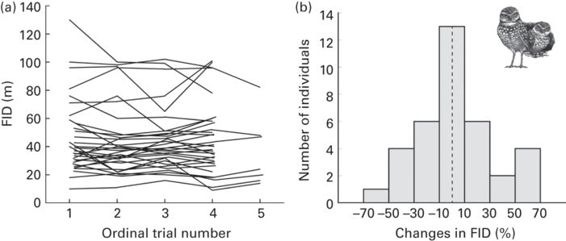
Figure 15.3
Consistency in the flight initiation distance (FID) of burrowing owls, Athene cunicularia. (a) Individual changes of FID across successive trials, for several individuals tested four or more times. (b) Percentage change in FID between the last and the first trial.
(Figure redrawn from Carrete & Tella 2010)
In the field cricket, Gryllus integer, individual differences in latency to become active after defensive freezing and in hiding time inside a refuge are consistent (Hedrick & Kortet 2012; Niemelä et al. 2012a). However, individual differences in boldness are consistent only in a novel environment (control) but not after being exposed to a predator, which suggests the existence of a context-specific behavioral syndrome (Niemelä et al. 2012b). Moreover, hiding time is repeatable across metamorphosis stages in females, but not in males, which become shyer with maturation, probably because of the risk associated with calling for mates. Also, individuals have longer freezing times when they are nymphs than when they are adults (Hedrick & Kortet 2012).
In hermit crabs, Coenobita clypeatus, latency to hide and emerge from their shells are highly consistent in several contexts (i.e., when crabs were inverted, or in an open field, or when subjected to a predator visual stimulus, or to an electric shock). Moreover, there are correlations between the hiding time after suffering an electric shock and both the latency to hide and hiding time after presenting a visual predator stimulus (Watanabe et al. 2012). Similar individual consistency in hiding or tonic immobility responses is also found, for example, in other hermit crab species, Pagurus bernhardus (Briffa et al. 2008), mealworm beetles, Tenebrio molitor (Krams et al. 2014), larval salamanders, Ambystoma barbouri (Sih et al. 2003), and even in sea anemones (Actinia equina; Briffa & Greenaway 2011; Condylactis gigantea; Hensley et al. 2012).
Taken together, these studies suggest that escape and hiding behavior may be consistent repeatable personality traits in many animals, at least in some contexts or during some ontogenetic states. In fact, the reported repeatability values for FID and hiding times are in many cases among the highest repeatabilities observed for behaviors (Réale et al. 2007; Bell et al. 2009).
15.2.2 Plasticity and habituation of the escape and hiding response
Despite demonstrable individual consistent differences, animals may also modify their behavior to varying environmental conditions (Dingemanse et al. 2010). This behavioral plasticity could be either similar among individuals, hence maintaining personality differences across an environmental gradient, or vary among individuals and situations (Nussey et al. 2007; Dingemanse et al. 2010; Figure 15.1). Several studies have revealed that both behavioral plasticity and consistency between situations often coexist. For example, larval salamanders, Ambystoma barbouri, spend more time out of their refuges in the absence of a fish predator than when it is present, but individual responses are correlated between the two situations (Sih et al. 2003). Similarly, hermit crabs, Pagurus bernhardus, show variation in average startle hiding behavior responses between several predator-cue treatments, indicating that there is behavioral plasticity between situations, but crabs also show a high individual consistency in the ranks of individual hiding times between situations (Briffa et al. 2008).
Habituation to predators is a special case of behavioral plasticity by which animals reduce their antipredator responses to a potential predatory stimulus through a process in which the stimulus ceases to be regarded as dangerous after repeated non-threatening exposures to it (e.g., Hemmi & Merkle 2009; Rodríguez-Prieto et al. 2010, 2011). Habituation to low-risk predators is important because it may have fitness consequences. For example, individual Iberian wall lizards, Podarcis hispanica, that habituate (i.e., decrease their FID) more readily to a frequent low-risk predatory stimulus (a human, that does not attack, but passes by close to the lizard) are able to increase their body condition more than lizards that habituate less (Rodríguez-Prieto et al. 2010; Figure 15.4).
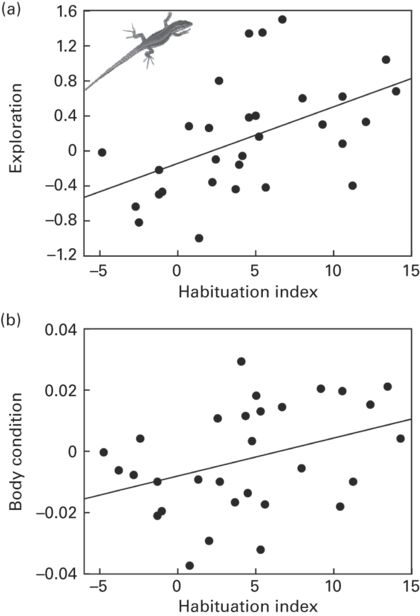
Figure 15.4
Inter-individual variation in habituation in Iberian wall lizards, Podarcis hispanica. Relationship between habituation index (higher values indicate a higher and quicker reduction of FID over the course of a six-day experiment with repeated approaches) and (a) exploration behavior (higher values indicate fast explorers) or (b) body condition change.
(Redrawn from Rodríguez-Prieto et al. 2010, 2011)
Several studies have used the progressive reduction in magnitude of FID in response to repeated predatory attacks as a measurable indicator of habituation (Lord et al. 2001; Runyan & Blumstein 2004; Magle et al. 2005; Hemmi & Merkle 2009; Rodríguez-Prieto et al. 2010, 2011; Carter et al. 2012b). For example, one study investigated the direct and indirect effects of boldness (estimated from refuge use, time being exposed, etc.), exploratory behavior in a novel environment and sociability (time spent hidden in a refuge with conspecific olfactory cues) on the inter-individual variability in habituation ability (i.e., reduction in FID) of Iberian wall lizards (Rodríguez-Prieto et al. 2011; Figure 15.4). Individual boldness was consistent across contexts, but it did not affect differences in habituation. However, exploration had a strong direct effect on habituation, with more exploratory individuals being able to habituate faster than less exploratory ones, probably because they could gather more information from novel stimuli and were able to assess risk better. Individual variation in habituation was also indirectly affected by sociability, probably because less social individuals that avoided refuges with conspecific cues, increased exposure to the predator and eventually habituated.
In contrast, repeated observation of escape behavior of different individual male rock agamas revealed a small, but significant, decline in FID with time, which suggests habituation to the observer’s approaches over the course of the study. However, individuals do not differ in the rate of habituation, which suggests that there are not inter-individual differences in behavioral plasticity (Carter et al. 2012a,b).
15.2.3 Correlations of escape and hiding behavior with other behavioral traits
Inter-individual variability in escape responses and hiding behavior from predators may be correlated with other personality traits such as activity level and exploration, thus providing strong evidence for a behavioral syndrome. For example, juvenile convict cichlids, Amatitlania nigrofasciata, that are more exploratory in a novel environment are slower to react to an attack by a simulated fish predator (Jones & Godin 2010). Similarly, great tits, Parus major, which are more willing to explore more of a novel environment, are also more likely to return to forage quicker after being startled (Van Oers et al. 2004b). And individual chaffinches, Fringilla coelebs, which are less active, are more likely to freeze than flee in response to a hawk attack and take longer to resume activity after the attack (Quinn & Cresswell 2005b). These syndromes could be explained by models of optimal escape behavior (Ydenberg & Dill 1986; Cooper & Frederick 2007b; see Chapter 2), with bolder, more exploratory individuals delaying their escape response and shortening hiding times to reduce the costs of missed foraging opportunities. Alternatively, consistent with the “perceptual limits hypothesis” (Quinn & Cresswell 2005a), individual prey differing in personality might have a different capacity to detect and react to predator attacks. Nevertheless, the latter may be tested in species that show alert responses before fleeing by looking at variation and consistency in alert distances.
Other studies have found a relationship between boldness toward predators and aggressive behavior toward conspecifics, although only in some populations of the same species but not in others (Huntingford 1976; Riechert & Hedrick 1993; Bell 2005). For example, individual breeding three-spined sticklebacks, Gasterosteus aculeatus, with higher levels of territorial aggression to intruders, are bolder toward a predator outside the breeding season (Huntingford 1976). In the spider Agelenopsis aperta, following a web disturbance that makes them hide, individuals that return more quickly to a foraging position at the funnel entrance of their webs are later more likely to win conspecific agonistic contests than spiders with longer return latencies (Riechert & Hedrick 1993). Male turtles, Mauremsy leprosa, that are bold in an antipredatory situation (i.e., with shorter hiding times inside the shell) avoid water with chemicals of unfamiliar competitor males, but not with chemicals of familiar males, whereas shy turtles avoid chemicals of both familiar and unfamiliar males (Ibañez et al. 2013). Bold individuals with respect to social interactions might also be bold under threatening situations because of high testosterone levels (Huntingford 1976; Tulley & Huntingford 1988). This may reflect a trade-off caused by the existence of behavioral correlations across functional contexts (Sih et al. 2003) because different traits have the same underlying physiological and genetic bases as pleiotropic effects of the same genes, which may not be necessarily adaptive in the context of predator avoidance. However, population comparisons and experimental work have shown that boldness and aggressiveness tend to covary in “high-predation” populations, where exposure to predation would generate this behavioral syndrome (Bell 2005; Dingemanse et al. 2007). One experimental study with sticklebacks, suggests that this relationship may be explained because while bolder individuals more willing to forage under predation risk are less likely to survive predation, those individuals that are more aggressive toward conspecifics are more likely to survive exposure to predators (Bell & Sih 2007), suggesting that this correlation between boldness and aggression may be adaptive in some environments.
However, boldness to predators is not necessarily correlated with boldness in other contexts. For example, in juvenile pumpkinseed sunfish, Lepomis gibbosus, individual consistent differences in responses to a threatening object do not correlate with individual consistent differences in response to a novel food source (Coleman & Wilson 1998). Similarly, dumpling squids, Euprymna tasmanica, that are bold in threat tests were not necessarily bold in feeding tests, and this lack of across-context correlations is observed across their entire life span (Sinn et al. 2008). These results suggest that shyness and boldness are context specific and may not exist as only a one-dimensional behavioral continuum, even within a single context. It is also possible that the lack of correlation tells us that responses to novel, non-threatening stimuli (neophobia) are not equivalent to responses to threatening predators or conspecifics.
15.3 Evolution of personalities and the optimal escape behavior
The economic or optimality models of escape and refuge use (Ydenberg & Dill 1986; Cooper & Frederick 2007a,b; see Chapter 2) do not explicitly consider inter-individual differences in personality. Such individual differences in boldness and other traits are a potentially important, but mostly neglected, source of variability in these antipredatory behaviors. The problem that arises is that if these models predict only one optimal escape response for each situation of risks and costs, we should initially expect that all individuals in the population will evolve to uniformly follow this optimal response. This is because individuals that fail to escape adequately from predators will incur higher predation risk, but individuals that have an exaggerated escape response will incur high costs in the form of missed foraging opportunities. Therefore natural selection would quickly favor individuals that show an optimal response (i.e., the one predicted by the models for each situation), and we should not expect to find inter-individual differences but rather a uniform escape strategy in all individuals of each population.
One possibility is that the optimality models are predicting what might be the average optimal response in each population, with observed differences being considered as the non-biologically important consequence of inaccurate measurements or small random non-adaptive variation around the adaptive mean (Wilson 1998). However, this does not exclude the fact that selection should eliminate those individuals with suboptimal responses. This would lead to uniform behavior after some time because survivors would be only those showing the single optimal response.
Other explanations for the existence of consistent inter-individual variability in personalities may be the existence of individual differences in state or life-history strategies (Dall et al. 2004; Wolf et al. 2007; Biro & Stamps 2008; Sih & Bell 2008; Luttberg & Sih 2010). Thus predictions of the models would apply separately to each individual because there is also variability in risk assessment (e.g., based on its escape abilities or life-history strategy) and costs of escape or refuge use (e.g., individuals with a better condition will have lower costs). These subtle differences in state or life-history strategies between individuals, which are not easy to measure, would explain the observed inter-individual variability in escape responses in apparently similar situations. We will expand and examine the empirical evidence for these hypotheses below.
15.3.1 State-dependent escape and hiding responses
Mathematical models suggest that among several state-dependent mechanisms, asset protection (i.e., individuals with more “assets” tend be more cautious) and starvation avoidance can explain short-term consistency in personality (Wolf et al. 2007, 2008), although not long-term stable personalities (McElreath et al. 2007). However, state-dependent safety (i.e., individuals with higher energy reserves, condition, or health are better at avoiding predators) can explain long-term stable differences in personality (Luttberg & Sih 2010). Therefore each individual would be optimizing its escape responses according to their own state and circumstances. In fact, many studies have shown that morphological and physiological factors, such as running speed, body weight, growth rates, tail loss, or health state, affect the escape responses and refuge use in many animals (Lindström et al. 2003; López et al. 2005; Biro & Stamps 2008; Jones et al. 2009; Cooper & Wilson 2010). Moreover, many of these state factors are permanent or of long-term duration in an individual, which will explain behavioral consistency through time, and their effects will be expressed across different contexts and correlated with other different behaviors, which in turn will result in behavioral syndromes (Dall et al. 2004). Therefore differences in the personality of escape behavior between otherwise apparently similar individuals can also be considered as adaptive.
Some studies support that inter-individual differences in escape behavior may reflect differences in state-dependent personalities between individuals. For example, when approached by skuas, Catharacta spp., unguarded old chinstrap penguin, Pygoscelis antarctica, chicks flee short distances and usually aggregate into a dense pack, which decreases individual predation risk. There is an apparently clear variability in the escape responses of different individual chicks under similar conditions. Some shy individuals flee sooner than others and even some bold individuals do not flee at all but remain still and face the approaching skua, threatening it with open bill and loud vocalizations (Martín et al. 2006). This variability might be related to the perceived vulnerability to predation of each individual chick. Simulated predatory attacks showed that inter-individual variability in escape behavior is partly explained by the social environment (i.e., the presence of some adults around that are able to deter a skua attack) and age (younger chicks do not escape because their parents are usually around to defend them). But in addition, chicks in relatively poor health, as indicated by their T-cell immune response, have greater FIDs and flee farther (Figure 15.5). Because health state affects general antipredatory behavior performance (Lindström et al. 2003), the ability of “poor” condition penguin chicks to defend themselves may be lower, forcing them to be shyer. Moreover, skuas and other avian predators might assess that bolder individual penguin chicks that escape relatively later or face the predator will be more difficult to capture, diverting their attacks to shyer individuals. Therefore bolder individuals might be signaling to predators their ability to escape and defend from a potential attack (see Chapter 10).
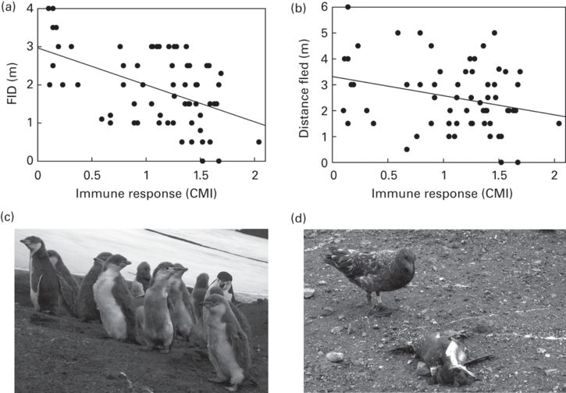
Figure 15.5
Inter-individual variability in state-dependent escape responses in unguarded chicks of chinstrap penguin, Pygoscelis antarctica. Relationships between the T-cell-mediated immune response (CMI) and (a) the flight initiation distance (FID) or (b) the distance fled of penguin chicks in response to a human predator attack simulating those naturally made by skuas, Catharacta spp. (c) Chinstrap penguins, Pygoscelis antarctica. (d) Skuas, Catharacta antarctica preying on a penguin chick.
(Figure redrawn from Martín et al. 2006; photographs by J. Martín)
Similarly, the escape response of incubating female eiders, Somateria mollissima, is modulated by interactions between stress responsiveness and individual quality (Seltmann et al. 2012). Younger females with higher stress responses (those with higher handling-induced corticosterone blood concentrations) have greater FIDs, whereas the opposite is found in the oldest females, which are considered of higher quality because they have more breeding experience. Furthermore, the effect of stress on FIDs is less marked in females in good body condition.
The sources of individual variation in antipredator behavior of a homogeneous group of adult male rock lizards, Iberolacerta cyreni, were examined in another study (Figure 15.6). In a laboratory setting, experimenters simulated repeated predatory attacks of low or high risk and analyzed activity levels, escape decisions and refuge use of lizards in both risk situations (López et al. 2005). Results showed consistent inter-individual differences in the magnitude of the antipredator responses under similar conditions of risk and costs. These individual differences in propensity to take risk could be caused by the existence of two consistent and independent shy-bold continua. The first shy-bold continuum describes the time spent hidden in the refuge after predatory attacks, with shy lizards hiding for longer. The second shy-bold continuum describes the propensity to make false-alarm flights and hide before an actual predator attack occurs (López et al. 2005). These shy-bold gradients can be related to the morphology and health state of each individual lizard: shy individuals that spend more time hidden in refuges have greater absolute body sizes, but relatively smaller head sizes (an index of a lower social dominance) and a worse body condition and immune system. However, shy individuals with a high propensity to hide when risk is low are those with smaller absolute body sizes, whereas their relative body size and the health state are not important (López et al. 2005). Interestingly, more dominant individuals were bolder. Similarly, territorial dominant males of the skink, Eulamprus heatwolei, were less likely than floater males to enter refuge when approached and returned to basking sites sooner after being attacked (Stapley & Keogh 2004). This may reflect the existence of a behavioral syndrome between boldness when using refuges and aggression toward conspecifics (see section 15.2.3 above).
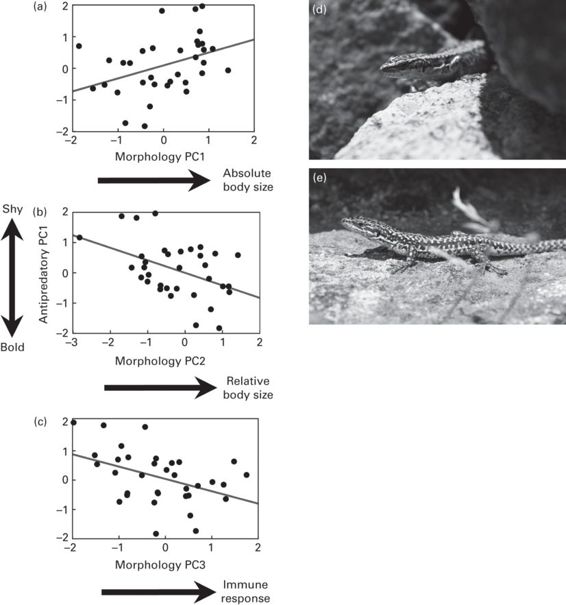
Figure 15.6
State-dependent hiding responses in adult male rock lizards, Iberolacerta cyreni. Relation between a shy-bold continuum (which describes a gradient from bold lizards that spent shorter times hidden in the refuge after predatory approaches to shy lizards with longer hiding times) and (a) absolute body size, (b) relative body size, and (c) immune response of lizards. (d, e) “Shy” and “bold” respectively, individual adult male rock lizards, Iberolacerta cyreni.
(Figure redrawn from López et al. 2005; photographs by J. Martín)
In yellow-bellied marmots, the cost of lost foraging opportunities interacts with personalities to determine hiding times inside refuges (Blumstein & Pelletier 2005; Figure 15.7). Bolder individuals that had shorter FIDs before hiding had subsequently shorter hiding times when food had been experimentally placed outside of their burrows before the attack than when no food was added. In contrast, shy individuals with longer FIDs, had subsequent longer hiding times if there were food present. These results might be explained by state-dependent responses whereby bold individuals were hungrier, and thus delayed escaping and hid for shorter periods of time. Alternatively, if bold individuals were less alarmed by the attack, they could be more able to estimate more accurately the costs of escape. However, both shy and bold marmots had similar hiding times in the no-food treatment.
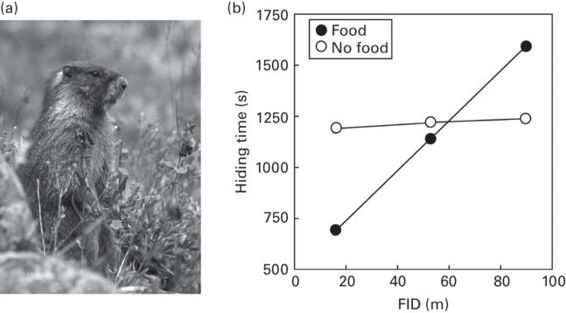
Figure 15.7
(a) A vigilant yellow-bellied marmot (Marmot flaviventris) monitoring an approaching predator.(b) The effect of FID (shy individuals have longer FIDs) on hiding time (mean ± SD) in their burrows by yellow-bellied marmots, Marmota flaviventris, as a function of whether supplementary food was presented or not.
(Photograph by B. Hulsey; figure redrawn from Blumstein & Pelletier 2005)
Results of these experiments suggest that the relative boldness of an individual when deciding escape or refuge use might be a function of state-dependent factors, such as its health and associated ability to evade predators. Moreover, all these characteristics can be considered as relatively fixed and phenotypically stable characteristics of an individual that might also be heritable. Persistence of personalities may emerge from positive feedback. For example, bolder individuals who are able to access food sooner, evade predators more easily, etc., and therefore maintain their higher states. In contrast, individuals in poor condition hide longer, lose opportunities to forage, and thus stay in a lower state.
15.3.2 Life history trade-offs when deciding escape responses
A meta-analysis (Smith & Blumstein 2008) suggests that the evolutionary mechanisms maintaining inter-individual variation in boldness may illustrate a trade-off between current and future reproduction (life history trade-offs hypothesis) with individual fitness varying depending on the context of the situation (Sih et al. 2004b; Wolf et al. 2007; Biro & Stamps 2008). Thus bolder individuals may have greater growth and reproductive success than shyer ones, but will suffer greater predation risk and have shorter life spans (productivity (growth)-mortality trade-off: Biro & Stamps 2008). In contrast, shy individuals may have reduced short-term reproductive success but will live longer by reducing predation risk, and therefore have the same overall life-time fitness as bold individuals (see models in Stamps 2007; Wolf et al. 2007). This trade-off may affect animal escape decisions.
Empirical evidence for the “life history trade-off hypothesis” (Wolf et al. 2007; Biro & Stamps 2008) with respect to escape responses is suggested from a field study of rock agamas (Carter et al. 2010). Bolder male agamas (those with consistently shorter FIDs) spend more time exposed in basking or moving and less time hidden than shyer males, suggesting the existence of a behavioral syndrome. These differences may have fitness consequences; bolder males benefit by having larger home ranges, which increase access to females and food, and higher feeding rates than shyer males. However, given that bolder males spent more time being conspicuous, they should suffer a higher predation risk, as inferred from a higher rate of tail loss (Carter et al. 2010). These observations suggest that bolder males may have higher reproductive success, but being bold may increase predation risk at a younger age, making their lifetime reproductive success equal to that of shyer males.
15.3.3 Negative frequency-dependent selection on escape responses
Although differences in state or life-history strategies may provide an explanation for personality differences (Dall et al. 2004; Biro & Stamps 2008; Sih & Bell 2008), there are many cases where individuals do not seem to differ in state, and it is not clear why a state-dependent response should be correlated with other behaviors in other contexts. Some alternative or complementary explanations to the evolution of personalities have been suggested, including negative frequency-dependent selection, spatio-temporal variations in the environment, or non-equilibrium dynamics, which could promote the coexistence of phenotypic variation in the population in stable frequencies (see review in Wolf et al. 2013). Nevertheless, these explanations account only for the existence of different personalities within a population but they do not explain why personality is consistent within an individual.
One study examined the role of negative frequency-dependent selection in inter-individual variations in antipredatory behavior of flocks of red knots, Calidris canutus, in response to a model sparrowhawk, Accipiter nisus (Mathot et al. 2011). Although flocks of knots differed in their escape behavior in relation to risk level, there was no between-individual variation in escape flight duration within flocks. In contrast, individuals differed in their degree of plasticity for the proportion of time spent vigilant across risk levels. These results support the prediction that traits with negative frequency-dependent payoffs (vigilance) may favor individual variation in plasticity (Wolf et al. 2008, 2013), while traits with positive frequency state-dependent payoffs (escape flights) may favor cohesiveness among group members.
15.4 Future directions
Although we have a relatively good understanding of the various trade-offs between avoiding predators and costs of the escape response and refuge use that an individual prey can face, we need to fully explain the causes of the inter-individual variation in escape strategies. However, most studies are based on single observations of each individual, while we need to track the same individuals in multiple occasions to test for consistency and repeatability of personalities in the different components of escape behavior and refuge use, and for the existence of plasticity and correlations between the different behaviors.
Also, to develop a comprehensive understanding of the evolutionary and ecological significance of personalities in escape responses, we need more integrative studies that follow a Tinbergian multidisciplinary approach (Sih et al. 2004b; Bell 2007). Thus we need to determine: (1) the proximate mechanisms (hormonal and neurological pathways, health state, etc.) underlying variation in escape responses; (2) the genetic basis of personalities, which may depend on both inherited and environmentally responsive genes, and may also suffer maternal effects (Bell 2009); (3) the factors that influence whether the escape responses and other behaviors (e.g., exploration, aggression) are decoupled or correlated as behavioral syndromes (Sih & Bell 2008); and (4) the fitness consequences of different escape strategies to test whether individuals with different personalities have similar success in evading predators, or whether, even if risk of predation is higher for some personality types, this is compensated for higher benefits in other contexts (foraging, reproduction, etc.) leading to similar lifetime fitness.
Finally, future theoretical models of optimal escape behavior and refuge use will have to incorporate relevant individuality into their structure/logic. Delaying escape or spending longer times hidden in a refuge than other individuals should not be considered as always maladaptive, but may simply represent alternative adaptive personalities, which may be understood when knowing the whole suite of behavioral responses and ecological demands of an animal.
References
Bell, A. M. (2005). Behavioral differences between individuals and populations of threespined stickleback. Journal of Evolutionary Biology, 18, 464-473.
Bell, A. M. (2007). Future directions in behavioural syndromes research. Proceedings of the Royal Society of London Series B, Biological Sciences, 274, 755-761.
Bell, A. M. (2009). Approaching the genomics of risk-taking behavior. Advances in Genetics, 68, 83-104.
Bell, A. M. & Sih, A. (2007). Exposure to predation generates personality in threespined sticklebacks (Gasterosteus aculeatus). Ecology Letters, 10, 828-834.
Bell, A. M., Hankison, S. J. & Laskowski, K. L. (2009). The repeatability of behaviour: a meta-analysis. Animal Behaviour, 77, 771-783.
Biro, P. A. & Stamps, J. A. (2008). Are animal personality traits linked to life-history productivity? Trends in Ecology and Evolution, 23, 361-368.
Blumstein, D. T. & Pelletier, D. (2005). Yellow-bellied marmot hiding time is sensitive to variation in costs. Canadian Journal of Zoology, 83, 363-367.
Boissy, A. (1995). Fear and fearfulness in animals. Quarterly Review of Biology, 70, 165-191.
Briffa, M. & Greenaway, J. (2011). High in situ repeatability of behaviour indicates animal personality in the beadlet anemone Actinia equina (Cnidaria). PLoS ONE, 6(7), e21963.
Briffa, M., Rundle, S. D. & Fryer, A. (2008). Comparing the strength of behavioural plasticity and consistency across situations: Animal personalities in the hermit crab Pagurus bernhardus. Proceedings of the Royal Society of London Series B, Biological Sciences, 275, 1305-1311.
Carere, C. & Maestipieri, D. (eds.) (2013). Animal Personalities. Behavior, Physiology and Evolution. Chicago, IL: University of Chicago Press.
Carrete, M. & Tella, J. L. (2010). Individual consistency in flight initiation distances in burrowing owls: A new hypothesis on disturbance induced habitat selection. Biology Letters, 6, 167-170.
Carter, A. J., Goldizen, A. W. & Tromp, S. A. (2010). Agamas exhibit behavioral syndromes: bolder males bask and feed more but may suffer higher predation. Behavioral Ecology, 21, 655-661.
Carter, A. J., Heinsohn, R., Goldizen, A. W. & Biro, P. A. (2012a). Boldness, trappability and sampling bias in wild lizards. Animal Behaviour, 83, 1051-1058.
Carter, A., Goldizen, A. & Heinsohn, R. (2012b). Personality and plasticity: Temporal behavioural reaction norms in a lizard, the Namibian rock agama. Animal Behaviour, 84, 471-477.
Coleman, K. & Wilson, D. S. (1998). Shyness and boldness in pumpkinseed sunfish: individual differences are context-specific. Animal Behaviour, 56, 927-936.
Cooper, W. E., Jr. (2009). Variation in escape behavior among individuals of the striped plateau lizard Sceloporus virgatus may reflect differences in boldness. Journal of Herpetology, 43, 495-502.
Cooper, W. E., Jr. (2011). Risk, escape from ambush, and hiding time in the lizard Sceloporus virgatus. Herpetologica, 68, 505-513.
Cooper, W. E., Jr. & Frederick, W. G. (2007a). Optimal flight initiation distance. Journal of Theoretical Biology, 244, 59-67
Cooper, W. E., Jr. & Frederick, W. G. (2007b). Optimal time to emerge from refuge. Biological Journal of The Linnean Society, 91, 375-382.
Cooper, W. E., Jr. & Wilson, D. S. (2010). Longer hiding time in refuge implies greater assessed risk after capture and autotomy in striped plateau lizards (Sceloporus virgatus). Herpetologica, 66, 425-431.
Cresswell, W., Quinn, J. L., Whittingham, M. J. & Butler, S. (2003). Good foragers can also be good at detecting predators. Proceedings of the Royal Society of London Series B, Biological Sciences, 270, 1069-1076.
Dall, S. R. X., Houston, A. I. & McNamara, J. M. (2004). The behavioural ecology of personality: Consistent individual differences from an adaptive perspective. Ecology Letters, 7, 734-739.
Dingemanse, N. J. & Dochtermann, N. A. (2013). Quantifying individual variation in behaviour: mixed effect modelling approaches. Journal of Animal Ecology, 82, 39-54.
Dingemanse, N. J. & Wolf, M. (2010) A review of recent models for adaptive personality differences. Philosophical Transactions of the Royal Society of London Series B, 365, 3947-3958.
Dingemanse, N. J., Thomas, D. K., Wright, J. et al. (2007). Behavioural syndromes differ predictably between twelve populations of three-spined stickleback. Journal of Animal Ecology, 76, 1128-1138.
Dingemanse, N. J., Kazem, A. J. M., Réale, D. & Wright, J. (2010). Behavioural reaction norms: Animal personality meets individual plasticity. Trends in Ecology and Evolution, 25, 81-89.
Garamszegi, L. Z. & Herczeg, G. (2012). Behavioural syndromes, syndrome deviation and the within- and between-individual components of phenotypic correlations: When reality does not meet statistics. Behavioral Ecology and Sociobiology, 66, 1651-1658.
Groothuis, T. G. G. & Carere, C. (2005). Avian personalities: characterization and epigenesis. Neuroscience & Biobehavioral Reviews, 29, 137-150.
Hedrick, A. V. & Kortet, R. (2006). Hiding behaviour in two cricket populations that differ in predation pressure. Animal Behaviour, 72, 1111-1118.
Hedrick, A. V. & Kortet, R. (2012). Sex differences in the repeatability of boldness over metamorphosis. Behavioral Ecology and Sociobiology, 66, 407-412.
Hemmi, J. & Merkle, T. (2009). High stimulus specificity characterizes anti-predator habituation under natural conditions. Proceedings of the Royal Society of London Series B, Biological Sciences, 276, 4381-4388.
Hensley, N. M., Cook, T. C., Lang, M., Petelle, M. B. & Blumstein, D. T. (2012). Personality and habitat segregation in giant sea anemones (Condylactis gigantea). Journal of Experimental Marine Biology and Ecology, 426-427, 1-4.
Huntingford, F. A. (1976). The relationship between anti-predator behaviour and aggression among conspecifics in the three-spined stickleback, Gasterosteus aculeatus. Animal Behaviour, 24, 245-260.
Ibáñez, A., Marzal, A., López, P. & Martín, J. (2013). Boldness and body size of male Spanish terrapins affect their responses to chemical cues of familiar and unfamiliar males. Behavioral Ecology and Sociobiology, 67, 541-548.
Jones, K. A. & Godin, J.-G. J. (2010). Are fast explorers slow reactors? Linking personality type and anti-predator behaviour. Proceedings of the Royal Society of London. Series B: Biological Sciences, 277, 625-632.
Jones, K. A., Krebs, J. R. & Whittingham, M. J. (2009). Heavier birds react faster to predators: Individual differences in the detection of stalking and ambush predators. Behavioral Ecology and Sociobiology, 63, 1319-1329.
Koolhaas, J. M., Korte, S. M., De Boer, S. F. et al. (1999). Coping styles in animals: Current status in behavior and stress-physiology. Neuroscience and Biobehavioral Reviews, 23, 925-935.
Krams, I., Kivleniece, I., Kuusik, A. et al. (2014). High repeatability of anti-predator responses and resting metabolic rate in a beetle. Journal of Insect Behavior, 27, 57-66.
Krause, J. & Godin, J.-G. J. (1996). Influence of prey foraging posture on flight behavior and predation risk: Predators take advantage of unwary prey. Behavioral Ecology, 7, 264-271.
Lindström, K. M., Van de Veen, I. T., Lagault, B. A. & Lundström, J. O. (2003). Activity and predator escape performance of common greenfinches Carduelis chloris infected with sindbis virus. Ardea, 91, 103-111.
López, P., Hawlena, D., Polo, V., Amo, L. & Martín, J. (2005). Sources of interindividual shy-bold variations in antipredatory behaviour of male Iberian rock-lizards. Animal Behaviour, 69, 1-9.
Lord, A., Waas, J. R., Innes, J. & Whittingham, M. J. (2001). Effects of human approaches to nests of northern New Zealand dotterels. Biological Conservation, 98, 233-240.
Luttbeg, B. & Sih, A. (2010). Risk, resources and state-dependent adaptive behavioural syndromes. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 365, 3977-3990.
Magle, S., Zhu, J. & Crooks, K. R. (2005). Behavioral responses to repeated human intrusion by black-tailed prairie dogs (Cynomys ludovicianus). Journal of Mammalogy, 86, 524-530.
Mathot, K. J., van den Hout, P. J., Piersma, T. et al. (2011). Disentangling the roles of frequency- vs. state-dependence in generating individual differences in behavioural plasticity. Ecology Letters, 14, 1254-1262.
Martín, J., de Neve, L., Fargallo, J. A., Polo, V. & Soler, M. (2006). Health-dependent vulnerability to predation affects escape responses of unguarded chinstrap penguin chicks. Behavioral Ecology and Sociobiology, 60, 778-784.
McElreath, R., Luttbeg, B., Fogarty, S. P., Brodin, T. & Sih, A. (2007). Communication arising: Evolution of animal personalities. Nature, 450, e5-e6.
Niemelä, P. T., Vainikka, A., Hedrick, A. V. & Kortet, R. (2012a). Integrating behaviour with life history: Boldness of the field cricket, Gryllus integer, during ontogeny. Functional Ecology, 26, 450-456.
Niemelä, P. T., DiRienzo, N. & Hedrick, A. V. (2012b). Predator-induced changes in the boldness of naïve field crickets, Gryllus integer, depends on behavioural type. Animal Behaviour, 84, 129-135.
Nussey, D. H., Wilson, A. J. & Brommer, J. E. (2007). The evolutionary ecology of individual phenotypic plasticity in wild populations. Journal of Evolutionary Biology, 20, 831-844.
Quinn, J. L. & Cresswell, W. (2005a). Escape response delays in wintering redshank, Tringa totanus, flocks: Perceptual limits and economic decisions. Animal Behaviour, 69, 1285-1292.
Quinn, J. L. & Cresswell, W. (2005b) Personality, anti-predation behaviour and behavioural plasticity in the chaffinch Fringilla coelebs. Behaviour, 142, 1377-1402.
Réale, D., Reader, S. M., Sol, D., McDougall, P. T. & Dingemanse, N. J. (2007). Integrating animal temperament within ecology and evolution. Biological Reviews, 82, 291-318.
Riechert, S. E. & Hedrick, A. V. (1993). A test for correlations among fitness-linked behavioural traits in the spider Agelenopsis aperta (Araneae, Agelinidae). Animal Behaviour, 46, 669-675.
Rodríguez-Prieto, I., Martín, J. & Fernández-Juricic, E. (2010). Habituation to low risk predators improves body condition in lizards. Behavioral Ecology and Sociobiology, 64, 1937-1945.
Rodríguez-Prieto, I., Martín, J. & Fernández-Juricic, E. (2011). Individual variation in behavioural plasticity: Direct and indirect effects of boldness, exploration and sociability on habituation to predators in lizards. Proceedings of the Royal Society of London. Series B: Biological Sciences, 278, 266-273.
Runyan, A. & Blumstein, D. T. (2004). Do individual differences influence flight initiation distance? Journal of Wildlife Management, 68, 1124-1129.
Seltmann, M. W., Ost, M., Jaatinen, K. et al. (2012). Stress responsiveness, age and body condition interactively affect flight initiation distance in breeding female eiders. Animal Behaviour, 84, 889-896.
Sih, A. & Bell, A. M. (2008). Insights from behavioral syndromes for behavioral ecology. Advances in the Study of Behavior, 38, 277-281.
Sih, A., Kats, L. B. & Maurer, E. E. (2003). Behavioural correlations across situations and the evolution of antipredator behaviour in a sunfish-salamander system. Animal Behaviour, 65, 29-44.
Sih, A., Bell, A. & Johnson, J. C. (2004a). Behavioral syndromes: an ecological and evolutionary overview. Trends in Ecology and Evolution, 19, 372-378.
Sih, A., Bell, A. M., Johnson, J. C. & Ziemba, R. E. (2004b). Behavioral syndromes: An integrative overview. Quarterly Review of Biology, 79, 241-277.
Sinn, D. L., Gosling, S. D. & Moltschaniwskyj, N. A. (2008). Development of shy/bold behaviour in squid: Context-specific phenotypes associated with developmental plasticity. Animal Behaviour, 75, 433-442.
Smith, B. R. & Blumstein, D. T. (2008). Fitness consequences of personality: A meta-analysis. Behavioral Ecology, 19, 448-455.
Stamps, J. A. (2007). Growth-mortality tradeoffs and “personality traits” in animals. Ecology Letters, 10, 355-363.
Stapley, J. & Keogh, J. S. (2004). Exploratory and antipredator behaviours differ between territorial and nonterritorial male lizards. Animal Behaviour, 68, 841-846.
Tulley, J. J. & Huntingford, F. A. (1988). Additional information on the relationship between intra-specific aggression and antipredator behaviour in the three-spined stickleback Gasterosteus aculeatus. Ethology, 78, 219-222.
Van Oers, K., De Jong, G., Drent, P. J. & Van Noordwijk, A. J. (2004a). Genetic correlations of avian personality traits: Correlated response to artificial selection. Behavior Genetics, 34, 611-619.
Van Oers, K., Drent, P. J., De Goede, P. & Van Noordwijk, A. J. (2004b). Realized heritability and repeatability of risk-taking behaviour in relation to avian personalities. Proceedings of the Royal Society of London Series B, Biological Sciences, 271, 65-73.
Watanabe, N. M., Stahlman, W. D., Blaisdell, A. P. et al. (2012). Quantifying personality: Different measures, different inferences. Behavioral Processes, 91, 133-140.
Wechsler, B. (1995). Coping and coping strategies: A behavioural view. Applied Animal Behaviour Science, 43, 123-134.
Wilson, D. S. (1998). Adaptive individual differences within single populations. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 353, 199-205.
Wolf, M., van Doorn, G. S., Leimar, O. & Weissing, F. J. (2007). Life-history trade-offs favour the evolution of animal personalities. Nature, 447, 581-584.
Wolf, M., van Doorn, G. S. & Weissing, F. J. (2008). Evolutionary emergence of responsive and unresponsive personalities. Proceedings of the National Academy of Sciences of the United States of America, 105, 15825-15830
Wolf, M., van Doorn, G. S., Leimar, O. & Weissing, F. J. (2013). The evolution of animal personalities. In Carere, C. & Maestipieri, D. (eds.) Animal Personalities. Behavior, Physiology and Evolution. Chicago: University of Chicago Press, pp. 252-275.
Ydenberg, R. C. & Dill, L. M. (1986). The economics of fleeing from predators. Advances in the Study of Behavior, 16, 229-249.