ESCAPING FROM PREDATORS An Integrative View of Escape Decisions (2015)
Part III Related behaviors and other factors influencing escape
14 Maternal and genetic effects on escape: a prospective review
Lesley T. Lancaster
14.1 Introduction
Despite our understanding that escape behaviors are often optimized under strong and chronic selection, few studies have examined how these behaviors are transmitted, and this lack of information impedes knowledge of how escape behaviors can evolve. In this chapter, I review studies that have identified potential mechanisms of escape behavior transmission, from estimates of heritability of escape behavior and other quantitative genetic parameters, to family-, population-, and species-level effects on escape that are suggestive of heritability and local adaptation, to specific maternal effect mechanisms that organize escape behavior. This review provides an overview of the current state of the field, with discusion of how escape behavior may become adaptively integrated phenotypically with other components of an animal’s biology, including morphology, life-history, and reproductive strategies. This chapter is intended as a foundation for further research directions and to identify the largest gaps in what is currently known. As predator-prey regimes are altered by human influences on the environment, it is becoming increasingly critical to determine whether and how prey animals will continue to optimize escape decisions. Understanding the evolvability of escape behavior is a critical component of this endeavor.
Predation imposes strong natural selection on individuals and populations of most animal species, resulting in evolution of antipredator traits that facilitate survival. Escape behavior is a particularly interesting component of any animal’s antipredator syndrome (i.e., the animal’s suite of traits that together facilitate survival against predation) because escape often represents the last line of defense. For example, most animals’ antipredator syndromes comprise multiple predator-avoidance traits, such as altered activity patterns, cryptic or warning coloration and behavior, vigilance, defensive nest construction, or chemical defenses. However, for many individuals, these predator avoidance mechanisms at some point ultimately fail to prevent the prey animal from being targeted by a predator for attack (Pinheiro 1996). At this point, escape behavior becomes a critical determinant of an animal’s survival, and the set of behaviors comprising “escape” might therefore be predicted to be refined by selection into a highly context specific and reliably produced sequence of actions.
There are a number of ways by which adaptive and effective escape behaviors may be organized at the individual level, including cultural transmission, learning by experience, innate expression of an entirely genetically determined behavior, and more complex, higher-order modes of transmission in which predator cues experienced by the parental generation are translated into developmental cues that influence offspring behavior (i.e., adaptive maternal effects). It is commonly assumed that some combination of these modes of transmission is responsible for the expression of most adaptive escape behaviors: the requirement that effective escape behaviors must be context specific suggests that phenotypic plasticity and incorporation of environmental cues may be important influences on the expression of such behaviors. On the other hand, escape behaviors have an immediate life or death outcome, affording limited opportunity for these behaviors to be shaped by learning or experience, particularly in solitary animals that cannot learn from witnessing attacks on conspecifics. For this reason, it is predicted or assumed that most escape behaviors are also based on a system of early-developmental or intergenerational transmission such as cultural (i.e., non-genetic, horizontal transmission), maternal (i.e., the components of vertical transmission between mothers and offspring besides direct genetic transmission of traits), or direct genetic mechanisms.
Despite the expectation that escape behaviors are modified under natural selection and controlled by adaptive genetic × environmental interactions (G×E), little is known about how these behaviors are transmitted. This is on one hand because escape behaviors are higher order traits that rely on underlying physiological (Chapter 13) and morphological (Chapter 11) characteristics, which themselves each have complex environmental and genetic bases. On the other hand, lack of a comprehensive understanding of how escape behavior is transmitted, organized, and refined by selection is likely due to the scarcity of studies that have tackled this topic. Studies focusing on the evolution of reaction norms for escape behavior are particularly rare. In this chapter, I review studies that have investigated either quantitative genetic or mechanistic bases of escape behavior transmission to synthesize what is currently known and point to fruitful future directions for research. To facilitate evaluation and future synthesis, many of the reviewed studies are summarized in Table 14.1.
Table 14.1 Summary of reviewed studies that examine the inheritance of escape behavior.
|
Taxon |
Behavior |
Effect |
Reference |
|
Estimated heritability |
|||
|
Scallops Argopecten purpuratus |
Latency to flee |
h2 = 0.45 ± 0.18 |
Brokordt (2012) |
|
Total number of claps (adductor muscle contractions) |
h2 = 0.51 ± 0.18 |
||
|
Clapping time |
h2 = 0.36 ± 0.17 |
||
|
Clapping rate |
h2 = 0.57 ± 0.18 |
||
|
Garter snakes Thamnophis ordinoides |
Distance fled |
h2 = 0.387 ± 0.169 - 0.798 ± 0.164 |
Brodie (1989, 1993) |
|
Number of reversals |
h2 = 0.225 ± 0.110 - 0.783 ± 0.184 |
||
|
Escape speed |
h2 = 0.419 ± 0.196 - 1.173 ± 0.163 |
||
|
Mamushi snakes Gloydius blomhoffii |
Distance fled |
h2 = 0.82 ± 0.47 (hunted site), 0.89 ± 0.50 (non-hunted site) |
Sasaki et al. (2009) |
|
Common fruit fly Drosophila melanogaster |
Distance fled |
h2 = 0.076 ± 0.011 (low line), 0.143 ± 0.021 (high line) |
Grant & Mettler (1969) |
|
Indirect evidence for heritability |
|||
|
Alpine swifts Apus melba |
Behavioral gradient from flee to attack |
Family-level effects (genetic or maternal) |
Bize et al. (2012) |
|
Garter snakes Thamnophis sirtalis |
Alert distance/flight initiation distance |
King (2002) |
|
|
Pea aphids Acyrthosiphon pisum |
Dropping behavior |
Andrade & Roitberg (1995) |
|
|
Guppies Poecilia reticulata |
Maneuverability/capture time |
Sire effects, non-maternal and likely genetic |
Evans et al. (2004) |
|
Field crickets Gryllus integer |
Hiding time |
Population-level effects in naïve progens, genetic or maternal |
Hedrick & Kortet (2006) |
|
Desert grass spiders Agelenopsis aperta |
Hiding time |
Population-level effects, likely genetic |
Riechert & Hedrick (1990) |
|
Trinidadian guppies Poecilia reticulata |
Capture time |
O’Steen et al. (2002) |
|
|
Streamside salamanders Ambystoma barbouri |
Stimulus to initiate flight |
Population-level and gene flow effects, likely genetic |
Storfer & Sih (1998) |
|
Checkered whiptail lizards Aspidoscelis tesselata |
Flight initiation distance, distance fled |
Population-level differences; maternal, genetic or learned |
Punzo (2007) |
|
Garter snakes Thamnophis sirtalis |
Proportion that flee |
Population-level effects, maternal-offspring correlation |
Placyk (2012) |
|
Common frog Rana temporaria |
Survival in predator presence |
Latitudinal cline |
Laurila et al. (2008) |
|
159 bird species |
Flight initiation distance |
Diaz et al. (2013) |
|
|
Sexual and unisexual whiptail lizards |
Flight initiation distance |
Species-level differences, correlation with sexual system |
Hotchkin & Riveroll (2005), see text for additional references |
|
Freshwater fish |
Flight initiation distance |
Species-level differences, correlation with body armor |
McLean & Godin (1989) |
|
Lizards (family: Cordylidae) |
Distance fled, proportion that enter refuge, escape speed, use of vertical surfaces in escape |
Losos et al. (2002) |
|
|
Maternal effects |
|||
|
Side-blotched lizards Uta stansburiana |
Vertical escape behaviors, evasive behaviors |
Egg size (VM/VP = 0.36) |
Lancaster et al. (2010) |
|
Red drum Sciaenops ocellatus |
Escape latency, flight initiation distance/alert distance, duration of escape |
Fatty acid content of eggs |
Fuiman & Ojanguren (2011) |
|
Painted dragons Ctenophorus pictus |
Latency to flee, assessment time |
Experimentally elevated yolk testosterone |
Tobler et al. (2012) |
|
Pied flycatchers Ficedula hypoleuca |
Proportion that flee |
Ruuskanen & Laaksonen (2010) |
|
|
Garter snakes Thamnophis elegans |
Reversal behaviors |
Maternal ecotype × experimentally elevated maternal corticosterone |
Robert et al. (2009) |
|
Common lizard Zootoca vivipara |
Latency to flee |
Experimentally elevated maternal corticosterone |
Meylan & Clobert (2004) |
|
Common lizard Zootoca vivipara |
Hiding time |
Experimentally elevated yolk corticosterone |
Uller & Olsson (2006) |
|
Grey partridge Perdix perdix |
Proportion that flee, crouching behavior |
Yolk carotenoids - no effect |
Cucco et al. (2006) |
|
Water pythons Liasis fuscus |
Stimulation to sustain flight, vertical behaviors |
Incubation temperature regime |
Shine et al. (1997) |
|
Keelback snakes Tropidonophis mairii |
Pauses during flight, proportion of individuals that enter refuge |
Webb et al. (2001) |
|
|
Oviparous scincid lizards (Bassiana duperreyi and Nannoscincus maccoyi) |
Reversals |
Incubation temperature regime |
Shine (1995) |
|
Three spine sticklebacks Gasterosteus aculeatus |
Proportion that orient to predator, survival |
Maternal experience |
McGhee et al. (2012) |
|
Fall crickets Gryllus pennsylvanicus |
Hiding time |
Storm & Lima (2010) |
|
|
Skinks Pseudemoia pagenstecheri |
Reversals |
Maternal condition |
Shine & Downes (1999) |
14.2 Empirical estimates of heritability
Because escape behavior has such a strong and direct link to survivorship with few opportunities for “practice,” effective escape strategies should have evolved high heritabilities. In practice, remarkably few studies have directly estimated the heritability of escape behaviors. Often, heritabilities of escape strategies are assessed as a component of personality (Chapter 15; Sinn et al. 2006). Conversely, other studies report significant heritabilities for traits that commonly underlie escape behavior, such as aspects of locomotor performance (Chapter 11), including sprint speeds and stamina (Garland 1988, Shaffer & Formanowicz 2000, Watkins & McPeek 2006, Blumstein et al. 2010). However, it is often unclear how selection acting on either higher (i.e., personality) or lower order (i.e., locomotor performance) traits will affect the evolutionary response of escape behavior itself. Studies that directly estimate escape behavior heritability are often performed in the laboratory, which minimizes stochastic environmental effects on behavior, but which may present an artificial picture of how animals actually behave in the wild. Two examples in which thorough quantitative genetic analyses have been carried out are included below as case studies. These are followed by a review of other studies providing less complete evidence of heritable escape behaviors in section 14.3.
14.2.1 Case study 1: Flight in scallops
Juvenile scallops (Argopecten purpuratus) exhibit significant heritabilities for escape behaviors in response to controlled episodes of contact with a sea star, a natural predator (Brokordt et al. 2012; Figure 14.1). Brokordt et al. estimated heritabilities using offspring of wild-caught (but laboratory-reared) parents that were produced in a full/half-sib breeding design. Escape behaviors measured included latency to flee following predator contact, number of claps performed by the scallop during escape (claps = contractions of the phasic adductor muscle, producing valve closings and openings), and the rate and duration of clapping, which determine the speed and distance of the scallop’s escape effort. Estimated heritabilities were high and significant for all behaviors (Table 14.1). In all cases, dam effects were near zero and non-significant, suggesting that escape is not strongly influenced by maternal effects in this system. Interestingly, when scallops were subjected to a repeat trial, heritabilities of escape behavior dropped considerably. Although behaviors were repeatable (R = 0.36-0.42), most were no longer significantly heritable when estimated from repeat performances. This suggests that naïve scallops exhibit innate and genetically based escape abilities that are subsequently influenced by experience or limiting energetic considerations (only five minutes rest time was allowed between trials).
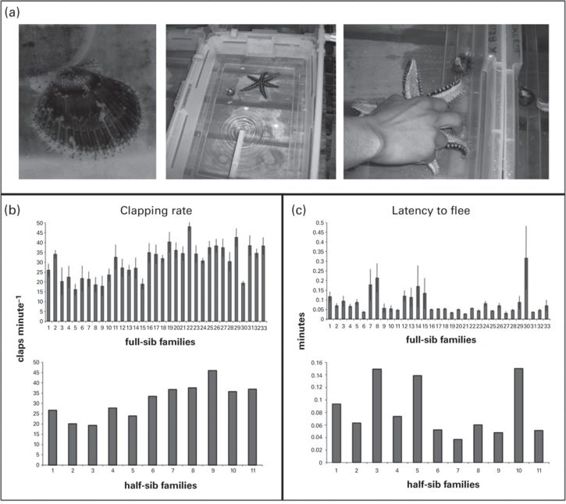
Figure 14.1
Heritable variation in scallop escape behavior (from Brokordt et al. 2012). (A) (left to right): Study organism, A. purpuratus. Escape trial arena. Escape trial in action (photo credits: Gabriela Núñez and Manuel Carmona). (B,C) Among-family variation in two of the measured escape behaviors, clapping rate (rate of contraction of phasic adductor muscle, providing an estimate of escape effort) and latency to flee following forced contact with a natural predator, a sea star.
(Figures reproduced with premission from K. Brokordt unpublished)
Escape behaviors in scallops were not only highly heritable, but also exhibited significant genetic correlations with each other: reaction time exhibited significant genetic correlations with clapping time (rG = 0.79 ± 0.21) and with number of claps (rG = - 0.74 ± 0.26), suggesting a genetic trade-off between fast reaction times and the ability to sustain the escape. Such a trade-off between speed and endurance has been commonly proposed (Garland 1988), but has not always been reported (Brodie 1993; Sorci et al. 1995). However, genetic correlations between different components of escape may be common, with some genetic correlations reflecting trade-offs and constraints, and others reflecting co-evolved suites of traits that work well together in an integrated escape strategy (see also section 14.2.2).
14.2.2 Case study 2: Flight in garter snakes
Garter snakes have become a model system for investigating the genetics of escape-related behavior (Arnold & Bennett 1984; Garland 1988) because they display a diversity of escape and defensive tactics combined with alternative antipredator markings (Jackson et al. 1976; Brodie 1989), and these traits each appear to be strongly genetically influenced. Brodie (1989, 1993) calculated heritabilities and genetic correlations among escape and other antipredator traits for individual garter snakes (Thamnophis ordinoides) based on laboratory behavioral trials of naïve, juvenile snakes and full-sib-based analyses of genetic parameters. Although full-sib analyses often conflate genetic and maternal effects, this conflation was partially removed by regressing behavior scores on potential indicators of maternal condition. Maternal effects that do not reflect maternal condition were not accounted for, however. As in Brokordt et al. (2012), significant and substantial heritabilities and genetic correlations were found for all escape and escape-related traits, including distance fled, number of reversals in direction of escape, and escape speed, with estimated heritabilities for escape behaviors ranging from 0.23 to 1.17. The highest and lowest estimates for each escape behavior’s heritability are reported in Table 14.1. Genetic correlations were reported between the number of reversals that a snake performed during flight and its dorsal patterning in two out of four populations (rG = - 0.328 ± 0.116 - rG = - 0.499 [s.e. not reported]). A significant genetic correlation between distance fled and speed was reported in one of the four populations (rG = 0.418 ± 0.111; Brodie 1993). In contrast with the scallop example above, faster garter snakes also exhibited more prolonged escapes.
Heritable variation in escape behavior of garter snakes is likely maintained under correlational selection favoring alternative escape syndromes: blotched snakes achieve high fitness by performing reversals to aid in crypsis whereas striped snakes survive best when escaping in a straight line to create an optical illusion of stasis (Brodie 1992; Figure 14.2A; see also section 14.5.1). Homogeneity of escape behavior × dorsal pattern G-matrices across diverged populations also suggests that these traits may be related by pleiotropy or linkage in addition to shared selection pressures (Brodie 1993) although much more work is needed to understand the genomic architecture of escape syndromes.
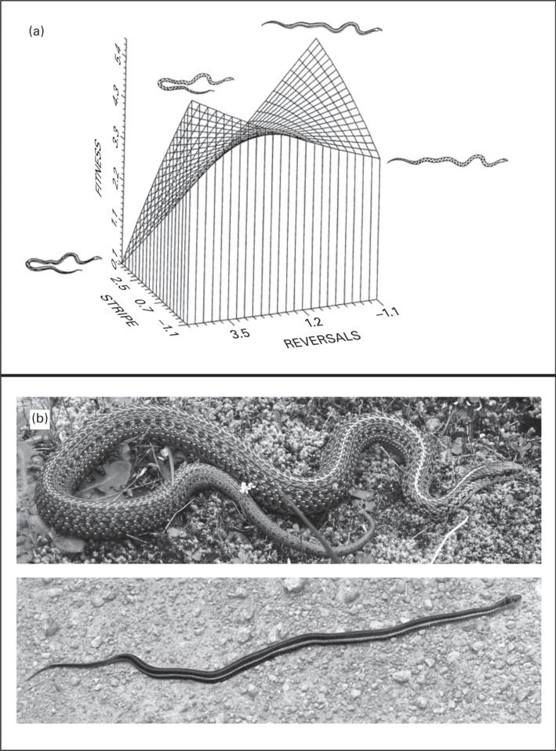
Figure 14.2
Genetic architecture of garter snake escape behavior (from Brodie 1989, 1993). (A) Correlational selection favors genetic correlations between dorsal patterning and escape behavior. Blotched snakes are selected to perform reversals, facilitating crypsis. Striped snakes are selected to escape in a straight line, facilitating an optical illusion of reduced speed. Figure reproduced with permission from Brodie (1992). (B) Escape behavior variation in T. ordinoides. Top panel: a blotched individual performing a reversal (photo by Gary Nafis). Bottom panel: a striped individual escapes in a straight line (photo by Jim Conrad).
14.3 Other evidence for genetic basis of escape behaviors
Although formal quantitative genetic studies of escape behavior are rare, many studies present at least partially convincing evidence for genetic bases of escape behavior. These cover a broad array of taxa, and can be grouped by the methodologies used to infer heritability, from parent-offspring or among-sib comparisons to artificial selection experiments to population- or species-level comparisons. Parent-offspring and full-sib comparisons establish family-level effects on escape behavior, although how this genetic component of phenotypic variation is distributed among maternal, additive, dominance, epistatic, and G × E effects remains to be determined. Artificial selection studies provide the additional advantage of supplying evidence for additive effects on escape behaviors, but these often cannot rule out maternal and grand-maternal effects, and are often conducted in systems so removed from the wild that it is difficult to apply their conclusions to natural systems. Finally, population- and species-level comparisons indicate larger scale trends in local adaptation and trade-offs that can inform future studies of the genetic architecture underpinning escape.
14.3.1 Family effects
Most commonly, familial effects on escape behavior have been detected via full-sib analysis or parent-offspring correlations under controlled conditions (either cross-fostering or common garden), and these effects may be attributed to either genetic or maternal/environmental effects of parents on progeny. For instance, adult, cross-fostered alpine swifts (Apus melba) behaved more like their biological parents than their foster parents in response to a simulated predator attack, suggesting either genetic or prenatal maternal effects on escape behavior (Bize et al. 2012). Sasaki et al. (2009) studied the repeatability and heritability of distance fled in response to human predators in the commercially valuable Japanese mamushi snakes (Gloydius blomhoffii), using full-sib comparisons that did not control for maternal effects. They found that flight distance exhibited significant family effects in both hunted and non-hunted populations (Table 14.1).
Alert distance/flight initiation distance (assessed as the distance at which the snake reacted to a simulated predator by either orienting or fleeing) showed a significant family effect in garter snakes (Thamnophis sirtalis), in full-sib comparisons in lab-reared progeny of wild-caught dams (King 2002). King also found significant family × time and family × time × testosterone interactions, suggesting that the ontogeny of escape behavior and the response of escape behavior to circulating steroid hormones are also either genetically based or subject to strong maternal effects. This study provides baseline evidence for a heritable set of environmentally modified reaction norms for escape behavior, a mechanism of inheritance that allows escape responses to be both reliably produced over multiple generations (i.e., stereotyped) and also suited to current conditions (i.e., individual or maternal experiences of predator regime, habitat, or individual variation in trade-offs between escape and other activities).
In one of the more convincing examples of genetic effects on escape behavior, Evans et al. (2004) tested for a genetic correlation between bright coloration and effective escape behavior in guppies (Poecilia reticulata) by artificially inseminating females with sperm from brightly or dully colored males with which the females had not had prior contact. Artificial insemination prevented females from using male coloration as a signal to trigger maternal effects, while testing the hypothesis that coloration is a reliable indicator of male genetic quality (Hill 1991). More brightly colored males sired offspring with better maneuvering ability during escape, and thus longer times to capture (by a human using a net), suggesting a genetic sire effect on escape behavior, and a genetic correlation between escape behavior and sexually selected coloration (Evans et al. 2004).
14.3.2 Evidence of heritability from artificially selected lines
A response to artificial selection over multiple generations in a controlled environment is likely sufficient to dispel all but the most persistent of confounding maternal effects, and this methodology has provided some solid evidence of genetic effects on escape behavior. For instance, parthenogenic pea aphids (Acyrthosiphon pisum) exhibit variation in their tendency to drop from a perch in response to alarm pheromone emitted by clone-mates. Andrade and Roitberg (1995) created three to four generation, divergently selected lines for this escape behavior in three clones of pea aphids, resulting in significant divergence in phenotype in the majority of cases.
In a related study, divergently selected lines of Drosophila melanogaster exhibited phenotypic divergence in distance fled upward through an I-maze away from vibrations imparted to the bottom of the maze (Grant & Mettler 1969). Escape behavior that continued to diverge over 18 generations without plateau, and also resulted in a correlated evolutionary response in assortative mating 'font-size:13.5pt;font-family:"inherit",serif; color:#5D6CEB'>Table 14.1). However, due to the highly artificial conditions of the behavioral trials, it is unclear how these results reflect true antipredator escape behaviors in a wild context.
14.3.3 Evidence for heritability and local adaptation from population comparisons
To infer local adaptation in escape behaviors, a number of investigators have examined the behaviors of lab-reared offspring of wild-caught mothers from populations subject to differing predation levels or regimes. Although these studies often cannot distinguish between genetic and maternal effects on offspring escape behaviors, it is possible in controlled rearing conditions to control for the effect of the offspring environment. Such controlled studies provide good evidence that at least some components of escape behaviors that vary among populations are organized before birth. Population comparisons represent the most abundant current evidence for heritability and local adaptation in escape behaviors.
Naïve, captive-reared offspring of wild-caught female field crickets (Gryllus integer) exhibited longer hiding times when their mothers had been captured from a population characterized by a greater diversity of predators than offspring from a population with fewer predator species (Hedrick & Kortet 2006). Subjects were obtained from two populations that differed in the number of predator species present and which were sufficiently geographically isolated to prevent high levels of gene flow, and escape behaviors were assessed in a controlled laboratory setting. A similar result for population level variation in hiding times was reported for lab-reared, F2 desert grass spiders Agelenopsis aperta from habitats characterized by differing avian predation intensities (Reichert & Hedrick 1990). Breeding to a second generation limits the influence of single-generation maternal effects, and provides strong evidence of genetic transmission and local adaptation in escape behavior. O’Steen et al. (2002), Punzo (2007), Placyk (2012), and Sasaki et al. (2009) have provided similar examples of population-level variation in escape behaviors in guppies, whiptail lizards, and snakes (Table 14.1).
Population-level comparisons are more convincing when couched in a formal population-genetics framework. Laboratory reared larval salamanders (Ambystoma barbouri) from populations with fish predators present required lower stimulus intensity (number of taps) to initiate flight than naïve larvae from populations in fishless habitats (Storfer & Sih 1998). Furthermore, the number of taps required to initiate escape depended on the degree of genetic isolation of the individual’s source population from a population not exposed to fish, indicating that gene flow constrains local adaptation in escape behaviors. Survival rates in the presence of a cichlid predator were also higher among individuals from populations more genetically isolated from populations not exposed to predation by fish. This example clearly indicates a genetic effect on escape and suggests a trade-off between escape and some other aspect of performance that is favored in the absence of predators.
Clinal gradients in escape behavior can also indirectly implicate local adaptation over large-scale gradients in selective regimes. Latitudinal gradients in escape behavior, corresponding to decreases in predator densities at high latitudes, offer suggestive evidence of adaptive genetic effects on escape behavior in frogs (Laurila et al. 2008) and birds (Díaz et al. 2013).
14.3.4 Species-level comparisons
Species-level comparisons offer evidence complementary to that obtained from population-level comparisons, providing insight into the evolution of escape behavior at macroevolutionary scales. Studies reviewed above suggest that the maintenance of adaptive differences in escape behavior among populations may depend on low gene flow (Storfer & Sih 1998), and, in turn, population-level differences in escape behavior may contribute to reproductive isolation among populations (Grant & Mettler 1969; Nakayama & Miyatake 2010). When an ecologically relevant trait such as escape behavior varies among populations and contributes to reproductive isolation between populations, this trait may facilitate or drive ecological speciation and adaptive radiations. The evidence for effects of divergently selected escape behavior on reproductive isolation among conspecific populations indicates that escape behavior may drive speciation dynamics in some systems. In other cases, escape behavior may exhibit phylogenetic and niche conservatism. Phylogenetic conservatism could indicate that escape behavior is developmentally or ecologically constrained, and strongly suggests a genetic basis for escape behavior and its correlations and trade-offs with other traits.
In a comparison of three congeneric species of whiptail lizards (Aspidoscelis spp.), two parthenogenic and one gonochoristic, species-level differences in flight initiation distance and other defensive behaviors were reported (Hotchkin & Riveroll 2005), consistent with some previous observations (Milstead 1957; Schall & Pianka 1980; Price 1992). This suggests that escape behavior might be affected by genetic changes that occur in the transition to parthenogenesis or subsequent loss of genetic variability (Schall & Pianka 1980; Paulissen 1998) although more evidence is needed to confirm the cause of variation in escape behavior across these species.
Flight initiation distance in four populations from three species of freshwater fish was inversely correlated with degree of body armor (McLean & Godin 1989). The authors argued that this correlation reflects an evolved trade-off between behavioral and morphological defense (see also Abrahams 1995). These studies rely on relatively few species comparisons and do not consider effects of phylogeny or divergence times.
In a phylogenetically informed comparison of morphological defense and escape behavior in 15 cordylid lizard species, more heavily armored species ran shorter distances (distance fled), entered refuges more frequently (proportion of individuals that enter refuge), ran more slowly, and were less likely to utilize vertical surfaces during flight (Losos et al. 2002). Armor was concluded to be part of an evolved, genetically based antipredator syndrome that involves altered escape behavior and habitat use. Alternatively, differences in escape behavior could be non-genetic and result from behavioral plasticity producing similar behaviors in the habitats used by armored species. In general, Losos et al. found that phylogenetic comparisons provided similar results to non-phylogenetic tests of escape behavior and correlations with morphology, indicating a high degree of evolutionary lability. More phylogenetic tests of escape behavior are required to complement population-genetic analyses and within-population heritability estimates, in order to better understand the genetic architecture and evolutionary potential of escape behavior.
14.4 Conclusions based on case studies and other evidence of escape heritability
The genetics of escape behavior is currently a neglected field. Very few formal quantitative analyses of escape behavior have been conducted. Even among those, maternal effects are not always effectively excluded (e.g., Brodie, 1993), and dominance and epistasis have not yet been addressed. The recent advancements in computationally intensive methods for quantitative genetic and genomic analysis within the last decade mean that such studies are now much more feasible. However, most studies to date provide only partial or equivocal evidence for heritability of escape behavior. Collectively, these studies strongly indicate that escape behavior has a genetic basis in a wide variety of prey taxa, often exhibits local adaptation, and may be associated with predictable antipredator syndromes (such as degree of morphological defense) or sexual/genetic systems (e.g., parthenogenesis, sexually selected coloration). As predicted, many of these studies indicate that escape behavior is likely to exhibit evolved plasticity and ontogenetic modification, and high evolutionary potential.
14.5 Maternal effects
Adaptive maternal effects are predicted to arise when a female reliably experiences one of several different environments, and the environment that she experiences also reliably predicts conditions her offspring are likely to face (Mousseau & Fox 1998). Because predator regimes often persist at intermediate ecological scales of space and time and/or exhibit cyclical dynamics (May 1972), escape behaviors are ideal candidates for adaptive maternal modification. Theoretically, maternal effects can reliably produce a range of adaptive behaviors in offspring according to local cues, in contrast to locally adapted behaviors with a strictly genetic basis, which suffer from slower evolutionary responses to changing predator regimes, and from gene flow from areas having different predation regimes (Storfer & Sih 1998). Furthermore, adaptive maternal effects are predicted to appropriately adjust offspring behaviors according to phase of a predator-prey density cycle.
Many of the studies on familial effects may be most plausibly interpreted as partially or substantially due to maternal effects, but convincing evidence is lacking. In contrast, studies that specifically address maternal effects on escape behavior do not commonly include a formal quantitative genetic analysis of the maternal contribution to phenotypic variation (VM). Instead, investigators of maternal effects on escape behavior have, to date, more often focused on behavioral and physiological mechanisms, including provisioning, incubation regimes, or other phenotypic interactions between mothers and their progeny that may or may not be adaptive. In these studies a combination of experimental manipulations and correlative approaches have been employed to identify specific effects of maternal traits on offspring escape behavior. Below I review a mechanism of maternal effects on escape behavior, with emphasis on a case study in which the role of maternal effects was assessed both mechanistically and in a quantitative genetics framework, and the maternal effect on escape behavior was placed in a broader context of an evolved character syndrome involving adaptive mating and antipredator strategies.
14.5.1 Case study: Egg size and alternative patterns of adaptive escape behavior in side-blotched lizards
Escape behaviors often exhibit adaptive variation within populations, where variation in escape behaviors prevents predators from matching their attack strategy to a single escape tactic (Schall & Pianka 1980). Furthermore, variable escape strategies are often most effective when paired with other traits that affect the success of each escape behavior in an integrated, multitrait antipredator strategy (e.g., adaptive correlations between particular escape behavior strategies and dorsal patterning or body size; Brodie 1989; DeWitt et al. 1999). Side-blotched lizards (Uta stansburiana) exhibit alternative throat colors that indicate alternative mating and reproductive tactics in each sex (Sinervo & Lively 1996). Throat color is correlated with egg size variation, with yellow-throated females laying larger eggs than orange-throated females (Sinervo et al. 2000). This species also exhibits continuous variation in dorsal patterning, with patterns ranging from longitudinal stripes to horizontal bars (Lancaster et al. 2007; Figure 14.3C). Lancaster et al. (2010) found that survival selection favored two alternative trait combinations. Yellow-throated lizards, which exhibit a non-territorial, sneaker strategy in males, survived best with a barred dorsal pattern combined with evasive escape behaviors such as reversals and zigzags during flight. This result confirms earlier results that blotched and barred patterns are often co-selected with cryptic and evasive escape behaviors in reptiles (Jackson et al. 1976; Brodie 1992; Figure 14.2A) because this combination of traits facilitates crypsis, particularly when escaping into camouflaging vegetation. Territorial orange-throated lizards, in contrast, survived best when exhibiting a striped dorsal pattern and vertical (e.g., jumping, climbing) behaviors. These findings corroborate previous reports that stripes facilitate quick escape by creating an optical illusion of reduced speed (Brodie, 1992; see also section 14.2.2).
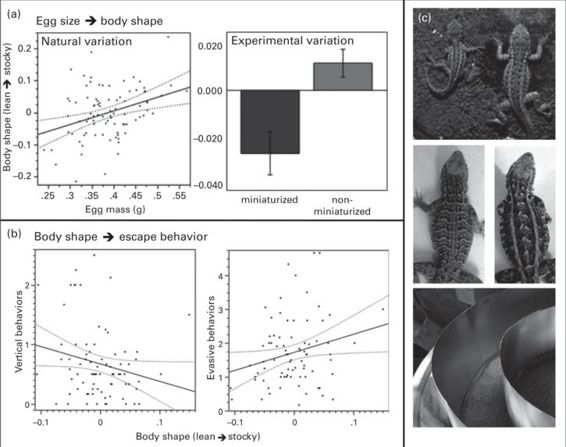
Figure 14.3
Effects of egg size on escape behavior (from Lancaster et al. 2010). (A) Egg size affects body shape in U. stansburiana. Left panel: regression of egg mass on body shape. Body shape, but not body size, effects persist throughout life (Figure 3C in original article, reproduced with permission). Right panel: experimental egg size manipulations also affect body shape. (B) Egg size effects on body shape direct alternative escape behaviors: leaner progeny are more likely to exhibit vertical behaviors (left panel), and stockier progeny are more likely to exhibit evasive behaviors (right panel). Experimental egg size manipulations produced similar effects on escape behavior (Figure 2C in original article, reproduced with permission). Data points in regressions represent full-sib means. (C) Top: effects of egg size manipulation on offspring body size and shape. Middle: examples of dorsal pattern variation; left = bars, right = stripes. Bottom: circular track for performing escape behavior trials
(Photo 14.3C top: by Barry Sinervo; middle and bottom: by Lesley Lancaster)
Adaptive combinations of such disparate traits as mating strategy, dorsal patterning, and escape behavior are rarely co-organized by a shared genetic pathway, and maternal effect plasticity may adaptively organize alternative character suites where linkage or pleiotropy are lacking (Lancaster et al. 2007, 2010). In side-blotched lizards, dorsal patterning and mating strategy are each heritable, but not genetically correlated with each other (Lancaster et al.2007), and dorsal patterning is also under maternal control via yolk estradiol (Lancaster et al. 2007). In the context of a half-sib crossing design, escape behaviors were measured ten times in succession in all parents and progeny. Measured escape behaviors included vertical behaviors (number of jumping + climbing attempts within each trial), evasive behaviors (zigzags + reversals + pauses), hiding behaviors (crouches + burying attempts), and total distance fled. Behaviors were repeatable, but heritability was low and non-significant for all behaviors. However, escape behaviors were significantly phenotypically correlated with both natural and experimental variation in body shape, with “lean” body types more likely to engage in vertical behavior, and “stocky” body types more likely to engage in evasive behavior (Lancaster et al. 2010; Figure 14.3B). Body shape is controlled by egg size in this and other lizard species (Figure 14.3A; Sinervo & Huey 1990), a maternal effect. Egg size itself has both a genetic and environmental basis (Sinervo et al. 2000; Lancaster et al. 2008). Body shape was experimentally adjusted via egg size manipulations (Sinervo & Huey 1990; Lancaster et al. 2010) to confirm that egg size itself, and not a correlated direct-genetic or maternal effect, was causally responsible for inducing the alternative escape behaviors (Figure 14.3A). Furthermore, escape behavior was confirmed to be under maternal influence in a quantitative genetic analysis, which indicated that the maternal component of phenotypic variation (VM, encompassing variance due to maternal effects and dominance) in progeny escape behavior was responsible for over a third of the total phenotypic variance (Table 14.1).
This study provides evidence that escape behavior may adaptively link alternative male and female reproductive strategies. In this study system, orange-throated males are territorial, inhabiting rock outcrops with good-quality nest sites. Orange-throated females tend to lay small eggs (resulting in lean hatchlings who perform vertical escape behaviors; these behaviors are beneficial in a three-dimensional, rocky habitat). Yellow-throated males are non-territorial sneakers that spend much of their time in the grass, and yellow-throated females tend to lay large eggs (resulting in stocky offspring that perform evasive behaviors that are beneficial in the between-territory vegetation). In the absence of an egg size effect on escape behaviors that are alternatively favored in habitats occupied by different mating strategy types, the genetic correlation between female egg size strategies and male mating strategies remains unexplained (egg size does not affect adult body size, but it does affect adult body shape; Lancaster et al. 2010). Similar correlations between mating and egg size strategies have been reported in other lizard species (Stapley & Keogh 2005), suggesting that escape behavior may commonly be responsible for observed correlations between alternative male mating strategies and female reproductive strategies.
14.5.2 Endocrine mechanisms of maternal effects on escape behavior
One of the most commonly studied ways that females can putatively affect the behavior of their progeny is by manipulating the prenatal endocrine environment, particularly by varying fetal exposure to steroid hormones. Prenatal glucocorticoid exposure (cortisol or corticosterone, depending on species) is a prime candidate for maternal organization of offspring escape behavior in vertebrates. This is because glucocorticoid levels, which function in vertebrate energy balance, are known to increase under stress and to affect behaviors requiring locomotion, such as activity level and dispersal (Dauphin-Villemant & Xavier 1987; De Fraipont et al. 2000). Glucocorticoids and other steroid hormones are easily transferred between mothers and developing offspring via the placenta or yolk of amniotic eggs (Hayward & Wingfield 2004). Therefore glucocorticoids provide a plausible, efficient mechanism by which mothers can communicate information about environmental stressors to offspring by using the same hormone that provides the cue to direct future offspring behavior (Uller & Olsson 2006), e.g., via known effects of glucocorticoids on energy distribution or muscle development (Chin et al. 2009).
Following this logic, several researchers have examined the effects of experimentally elevated maternal glucocorticoid levels on offspring escape behavior, mainly using reptilian study systems. Juvenile Lacerta (Zootoca) vivipara lizards from corticosterone-treated mothers exhibited increased latency to flee (required more stimulation to run) in comparison with offspring of control females (Meylan & Clobert 2004). Also in Z. vivipara, hatchlings from eggs that were directly injected with corticosterone exhibited significantly longer hiding times than control individuals (Uller & Olsson 2006). In the garter snake Thamnophis elegans, the effects of maternal corticosterone on escape behavior depended on maternal ecotype (Robert et al. 2009); specifically, experimentally elevated maternal corticosterone resulted in a reduced frequency of reversals in the “lakeshore” ecotype, which is characterized by a blotched dorsal pattern and frequent reversals. However, maternal corticosterone treatment had no effect on this behavior in the “mountain meadow” ecotype, which is characterized by stripes and few reversals. Ecotypes of T. elegans are correlated with alternative life-history strategies, and divergent naturally occurring corticosterone blood titers, suggesting that maternal effects on escape behavior may be closely tied to alternative life-history strategies (in addition to alternative mating and reproductive strategies; see sections 14.3 and 14.5.1). In each of these studies on maternal effects of glucocorticoids in reptiles, prenatal exposure to corticosterone was correlated with escape strategies involving lower energetic expenditure than in untreated controls.
Other steroid hormones have been investigated for potential maternal effects on escape behavior. Experimental prenatal exposure to androgens (testosterone injected into egg yolks) resulted in shorter assessment time and latency to flee in the painted dragon (Ctenophorus pictus), potentially indirectly via an effect on digit ratio (i.e., the relative lengths of individual digits on a single limb, which can affect locomotor performance; Tobler et al. 2011, 2012). Contrary to predicted effects of androgens on masculinized morphology and higher risk taking, experimentally elevated yolk testosterone in these studies resulted in a more feminized pattern of higher 3D:4D digit length ratios (ratio of the third to fourth digit length on a single limb), and a higher 3D:4D ratio was correlated with reduced risk-taking in escape behavior. In contrast, experimentally elevated yolk androgens (testosterone and androstenedione) resulted in the predicted pattern of increasingly risky escape behaviors in male pied flycatchers (Ficedula hypoleuca), which exhibited a decreased tendency to flee (~proportion that flee) (Ruuskanen & Laaksonen 2010).
The mechanism by which females increase offspring prenatal exposure to androgens in response to environmental cues is unknown, and likely involves a more complex physiological basis than glucocorticoid-based maternal effects, which can be transmitted as a passive response to maternal stress. Once androgens have been transferred to offspring, their effects on escape behavior are predicted to arise as downstream organizational effects on brain development, leading to increased overall levels of risk taking and aggressiveness (Partecke & Schwabl 2008). However, as Tobler et al. (2012) demonstrate, the effects of androgens on escape are often unpredictable and may have a more complex basis, including the aromatization of testosterone to estradiol by developing progeny (Crews et al. 1995). The potential effects of other (steroidal and non-steroidal) maternally derived hormones on escape behavior are currently unknown, with the exception of some preliminary evidence for no effect of maternal estradiol (Lancaster et al. 2010).
14.5.3 Other maternal provisioning effects on escape behavior
In addition to hormones, females provision offspring with varying quantities and qualities of many other nutrients and chemical cues that may have adaptive or incidental effects on offspring escape behavior. For instance, egg size influences escape behavior in side-blotched lizards via allometric body shape effects (section 14.5.1). Egg size is likely to commonly affect escape behavior in other species via its effect on offspring size because escape decisions are often size dependent (Wahle 1992, Shine et al. 1997, DeWitt et al. 1999). In addition to quantity of maternal provisioning, quality of provisioning may affect escape behaviors. The long-chain fatty acid content of eggs was correlated with offspring escape behaviors in red drum fish (Sciaenops ocellatus) although egg size itself had no effect (Fuiman & Ojanguren 2011). Long-chain fatty acids are usually associated with a high-quality diet and are important for nervous system function in vertebrates (Tocher 2003). Of 33 fatty acids tested, ten had significant effects on escape behavior. Effects were complex: some behaviors were influenced by multiple fatty acids, others by a single fatty acid type. In some cases, different fatty acids had opposing effects on the same escape variable. Understanding the generality and mechanisms of these effects will require further investigation.
Maternal investment in carotenoids represents an additional route by which females might increase offspring performance by increasing egg quality. Carotenoids are important in preventing damage due to oxidative stress and, like long-chain fatty acids, are obtained from high-quality diets (Sies 1997). Carotenoids are commonly invested in eggs (and are responsible for yellow yolk pigmentation), resulting in increased egg quality (Blount et al. 2002). However, carotenoid supplementation of laying females did not affect offspring escape behavior in the grey partridge, Perdix perdix (Cucco et al. 2006).
The putative link between compositional quality of eggs and escape behavior probably reflects an effect of provisioning on the overall condition and quality of offspring, with well-provisioned offspring better able to perform energetically costly or neurologically complex escape behaviors. Whether the quality of prenatal provisioning is in general an important contributor to effective escape performance is unknown. Furthermore, most studies of maternal provisioning effects have been conducted in egg-laying species, where provisioning is more easily quantified and characterized. In placental animals, the time interval available for prenatal provisioning is often more prolonged than in oviparous species. Therefore variation in the quantity and quality of resources provided by placental dams may reflect a broader range of conditions experienced by the mother. Although prenatal provisioning is more difficult to assess in placental species, such a study would provide a valuable comparison to existing information on egg-laying species.
14.5.4 Incubation regimes and maternal care
Incubation conditions, with or without maternal care, can affect escape behavior in reptiles. Nest site choice and incubation behaviors can introduce wide variation in the temperatures experienced by developing offspring, and incubation temperature has major effects on hatching time, offspring size, shape, sex, and behavior (Shine & Harlow 1996). Maternal choice of incubation temperature may be adaptively influenced by environmental cues, including predator density or predator type. Under natural selection, incubation temperature may be incorporated as a cue for offspring to develop appropriate escape strategies in response to the environment experienced by their mothers. Alternatively, variation in incubation temperature may have neutral or maladaptive effects on escape behavior if developmental conditions are suboptimal for neurological and musculoskeletal development.
Shine et al. (1997) investigated the effects of incubation regime on the amount of stimulation required by neonatal water pythons (Liasis fuscus) to sustain flight from a human predator through the water. Females exhibited alternative incubation strategies, choosing either to lay in varanid lizard burrows with hot, stable temperatures not requiring further attendance or to lay eggs in tree root boles having lower, more variable temperatures that require maternal attendance and shivering thermogenesis to maintain adequate incubation temperatures. Lab-reared hatchlings from eggs incubated in thermal regimes simulating (hot, stable) varanid burrow nests required more stimulation to sustain flight than hatchlings reared in conditions simulating (cooler, variable) tree bole nests, with or without maternal attendance. In contrast, hatchlings reared under root bole nest conditions, but without maternal attendance, exhibited increased vertical behaviors.
Incubation regimes reflecting natural variation in burrow depth affected escape behavior in a tropical natricine snake, the keelback, Tropidonophis mairii (Webb et al. 2001). Constant temperatures found in deep nests resulted in more pauses during flight, but a decreased tendency to hide, in comparison to the behavior of hatchlings incubated at the variable temperatures of shallow nests. The fitness consequences of offspring escape behaviors resulting from variation in incubation regimes, and whether these putative effects on offspring fitness have influenced the evolution of maternal incubation decisions, are unknown.
Shine (1995) investigated effects of incubation temperatures that mimic external nests vs. maternal body temperature in two oviparous scincid lizard species that are members of lineages in which viviparity has evolved. He found that cooler, nest-equivalent incubation temperatures resulted in hatchlings displaying more frequent reversals and defensive behaviors during flight. However, it is unknown whether the prenatal effects of maternal body temperature on offspring escape behavior are beneficial, or if they are involved in selection for viviparity.
14.5.5 Maternal environment and condition effects with unknown mechanisms
Several effects of maternal environment or condition on offspring escape behavior have been identified, but operate by unknown mechanisms. In a clear-cut example of a maladaptive maternal effect based on maternal experience, female three-spine sticklebacks that were exposed to predators produced offspring that were less likely to orient to the same predator type, and were therefore more likely to be eaten (McGhee et al. 2012). Conversely, female fall crickets (Gryllus pennsylvanicus) that were exposed to predators while gravid produced offspring with longer hiding times, which led to higher survival rates in the presence of predators (Storm & Lima 2010).
Maternal condition can also affect offspring escape behavior. For example, in the viviparous scincid lizard Pseudemoia pagenstecheri, offspring from females maintained with restricted food intake were more likely to perform reversals during escape, a behavior that was never seen in offspring from well-fed mothers (Shine & Downes 1999). The adaptive significance of this maternal effect is unknown.
14.5.6 Conclusions about maternal effects
Maternal effects on escape behavior have been reported in diverse taxa, albeit primarily in oviparous vertebrates. They operate by a wide variety of mechanisms, from effects of maternal behavior, experience, and condition to specific physiological effects of temperature and maternal provisioning. More studies are needed addressing the fitness effects of maternal effects on escape behavior in order to better discern adaptive maternal effects from maladaptive or neutral consequences of suboptimal prenatal and rearing conditions. More work is needed on viviparous vertebrates and invertebrates to complement existing studies, and to understand the generality of and interactions among different potential maternal effect mechanisms.
14.6 General conclusions
Evidence for a genetic basis or other prenatal organization of escape behavior has accumulated for diverse prey (Figure 14.4; Table 14.1) although data on invertebrates and mammals are sparse or lacking. One of the more interesting generalizations from findings to date is that escape behavior is often correlated with other aspects of an organism’s biology, including mating and reproductive strategies, life history, sexually selected coloration, genetic systems, and morphology. These correlations suggest that escape behavior is often highly integrated with other aspects of an organism’s biology, and that its heritability and mechanisms of transmission are strongly influenced and constrained by other, often non-predation related, traits. That escape behavior is often highly integrated with other traits also indicates its potential for use as an easily assessed indicator of the organism’s overall behavioral syndrome.
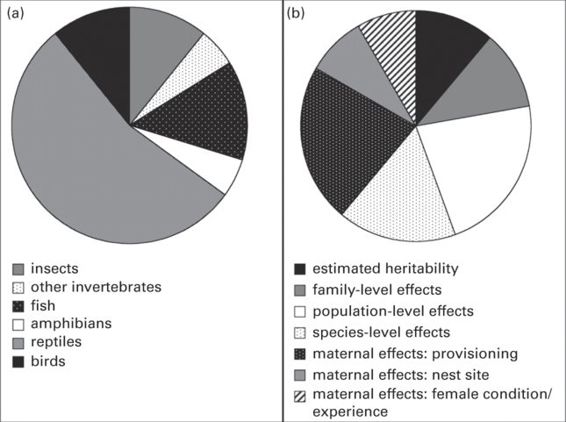
Figure 14.4
Summary of the current representation in the literature of studies examining the transmission of escape behavior, where each wedge represents the proportion of total papers reviewed: (A) by taxon; and (B) by mechanism. Studies were identified by searching Web of Science for combinations of the keywords and phrases: “maternal effect/s,” “escape behavior/our,” “antipredator behavior/our,” “heritable/ility,” “selection,” and “genetic/s,” and searching within cited articles and articles that cite keyword search results.
The literature reviewed here also testifies to the breadth of genetic, maternal, and environmental mechanisms that can affect various components of escape behavior. Overall, it confirms that escape behavior is subject to a wide range of influences affecting development, reflecting the dependence of variation in neurological and musculoskeletal development on the concerted action of a wide variety of underpinning genetic and physiological mechanisms. Because escape behavior is complex, many influences can potentially affect its effectiveness and/or adaptively direct its development. Further comparative studies are needed to determine how these trait associations evolve, and how escape is developmentally or ecologically constrained.
More research is needed to determine the generality of each of these mechanisms, to understand the genetic architecture of escape behavior evolution and to identify the evolutionary potential of escape behavior given correlations and trade-offs with other traits. Investigating the role of escape behavior in shaping macroevolutionary processes of speciation, facilitating niche evolution, and interacting with evolving sexual and genetic systems may prove fruitful. Further research is also needed to test for G × E interactions and specific genetic mechanisms that facilitate adaptive plasticity of escape behavior. Modern molecular ecological methods for identification of quantitative-trait loci (QTL), patterns of genome-wide association (GWA), and loci for adaptive traits in a landscape genetic framework are likely to be useful in determining maternal and genetic effects on escape behavior.
References
Abrahams, M. V. (1995). The interaction between antipredator behaviour and antipredator morphology: Experiments with fathead minnows and brook sticklebacks. Canadian Journal of Zoology, 73, 2209-2215.
Andrade, M. C. B. & Roitberg, B. D. (1995). Rapid response to intraclonal selection in the pea aphid (Acyrthosiphon pisum). Evolutionary Ecology, 9, 397-410.
Arnold, S. J. & Bennett, A. F. (1984). Behavioral variation in natural populations. 3. Antipredator displays in the garter snake Thamnophis radix. Animal Behaviour, 32, 1108-1118.
Bize, P., Diaz, C. & Lindstrom, J. (2012). Experimental evidence that adult antipredator behaviour is heritable and not influenced by behavioural copying in a wild bird. Proceedings of the Royal Society B-Biological Sciences, 279, 1380-1388.
Blount, J. D., Surai, P. F., Nager, R. G., et al. (2002). Carotenoids and egg quality in the lesser black-backed gull Larus fuscus: A supplemental feeding study of maternal effects. Proceedings of the Royal Society B-Biological Sciences, 269, 29-36.
Blumstein, D. T., Lea, A. J., Olson, L. E. & Martin , J. G. A. (2010). Heritability of anti-predatory traits: Vigilance and locomotor performance in marmots. Journal of Evolutionary Biology, 23, 879-887.
Brodie, E. D. (1989). Genetic correlations between morphology and antipredator behavior in natural populations of the garter snake Thamnophis ordinoides. Nature, 342, 542-543.
Brodie, E. D. (1992). Correlational selection for color pattern and antipredator behavior in the garter snake Thamnophis ordinoides. Evolution, 46, 1284-1298.
Brodie, E. D. (1993). Homogeneity of the genetic variance-covariance matrix for antipredator traits in 2 natural populations of the garter snake Thamnophis ordinoides. Evolution, 47, 844-854.
Brokordt, K., Farias, W., Lhorente, J. P. & Winkler, F. (2012). Heritability and genetic correlations of escape behaviours in juvenile scallop Argopecten purpuratus. Animal Behaviour, 84, 479-484.
Chin, E. H., Love, O. P., Verspoor, J. J., et al. (2009). Juveniles exposed to embryonic corticosterone have enhanced flight performance. Proceedings of the Royal Society B-Biological Sciences, 276, 499-505.
Crews, D., Cantu, A. R., Bergeron, J. M. & Rhen, T. (1995). The relative effectiveness of androstenedione, testosterone, and estrone, precursors to estradiol, in sex reversal in the red-eared slider (Trachemys scripta), a turtle with temperature-dependent sex determination. General and Comparative Endocrinology, 100, 119-127.
Cucco, M., Guasco, B., Malacarne, G. & Ottonelli, R. (2006). Effects of beta-carotene supplementation on chick growth, immune status and behaviour in the grey partridge, Perdix perdix. Behavioural Processes, 73, 325-332.
Dauphin-Villemant, C. & Xavier, F. (1987). Nychthemeral variations of plasma corticosteroids in captive Lacerta vivipara Jacquin: Influence of stress and reproductive state. General and Comparative Endocrinology, 67, 292-302.
De Fraipont, M., Clobert, J., John-Alder, H. & Meylan, S. (2000). Increased pre-natal maternal corticosterone promotes philopatry of offspring in common lizards Lacerta vivipara. Journal of Animal Ecology, 69, 404-413.
DeWitt, T. J., Sih, A. & Hucko, J. A. (1999). Trait compensation and cospecialization in a freshwater snail: Size, shape and antipredator behaviour. Animal Behaviour, 58, 397-407.
Díaz, M., Møller, A. P., Flensted-Jensen, E., et al. (2013). The geography of fear: A latitudinal gradient in anti-predator escape distances of birds across Europe. Plos One, 8, 7.
Evans, J. P., Kelley, J. L., Bisazza, A., Finazzo, E. & Pilastro, A. (2004). Sire attractiveness influences offspring performance in guppies. Proceedings of the Royal Society B-Biological Sciences, 271, 2035-2042.
Fuiman, L. A. & Ojanguren, A. F. (2011). Fatty acid content of eggs determines antipredator performance of fish larvae. Journal of Experimental Marine Biology and Ecology, 407, 155-165.
Garland, T. (1988). Genetic basis of activity metabolism - 1. Inheritance of speed, stamina, and antipredator displays in the garter snake Thamnophis sirtalis. Evolution, 42, 335-350.
Grant, B. & Mettler, L. E. (1969). Disruptive and stabilizing selection on the escape behavior of Drosophila melanogaster. Genetics, 62, 625-637.
Hayward, L. S. & Wingfield, J. C. (2004). Maternal corticosterone is transferred to avian yolk and may alter offspring growth and adult phenotype. General and Comparative Endocrinology, 135, 365-371.
Hedrick, A. V. & Kortet, R. (2006). Hiding behaviour in two cricket populations that differ in predation pressure. Animal Behaviour, 72, 1111-1118.
Hill, G. E. (1991). Plumage coloration is a sexually selected indicator of male quality. Nature, 350, 337-339.
Hotchkin, P. & Riveroll, H. (2005). Comparative escape behavior of chihuahuan desert parthenogenetic and gonochoristic whiptail lizards. Southwestern Naturalist, 50, 172-177.
Jackson, J. F., Ingram, W. & Campbell, H. W. (1976). Dorsal pigmentation pattern of snakes as an antipredator strategy: A multivariate approach. American Naturalist, 110, 1029-1053.
King, R. B. (2002). Family, sex and testosterone effects on garter snake behavior. Animal Behaviour, 64, 345-359.
Lancaster, L. T., McAdam, A. G., Wingfield, J. C. & Sinervo, B. R. (2007). Adaptive social and maternal induction of antipredator dorsal patterns in a lizard with alternative social strategies. Ecology Letters, 10, 798-808.
Lancaster, L. T., Hazard, L. C., Clobert, J. & Sinervo, B. R. (2008). Corticosterone manipulation reveals differences in hierarchical organization of multidimensional reproductive trade-offs in r-strategist and K-strategist females. Journal of Evolutionary Biology, 21, 556-565.
Lancaster, L. T., McAdam, A. G. & Sinervo, B. (2010). Maternal adjustment of egg size organizes alternative escape behaviors, promoting adaptive phenotypic integration. Evolution, 64, 1607-1621.
Laurila, A., Lindgren, B. & Laugen, A. T. (2008). Antipredator defenses along a latitudinal gradient in Rana temporaria. Ecology, 89, 1399-1413.
Losos, J. B., Mouton, P. L. N., Bickel, R., Cornelius, I. & Ruddock, L. (2002). The effect of body armature on escape behaviour in cordylid lizards. Animal Behaviour, 64, 313-321.
May, R. M. (1972). Limit cycles in predator-prey communities. Science, 177, 900-902.
McGhee, K. E., Pintor, L. M., Suhr, E. L. & Bell, A. M. (2012). Maternal exposure to predation risk decreases offspring antipredator behaviour and survival in threespined stickleback. Functional Ecology, 26, 932-940.
McLean, E. B. & Godin, J. G. J. (1989). Distance to cover and fleeing from predators in fish with different amounts of defensive armor. Oikos, 55, 281-290.
Meylan, S. & Clobert, J. (2004). Maternal effects on offspring locomotion: Influence of density and corticosterone elevation in the lizard Lacerta vivipara. Physiological and Biochemical Zoology, 77, 450-458.
Milstead, W. W. (1957). Observations on the natural history of four species of whiptail lizard, Cnemidophorus (Sauria, Teiidae) in Trans-Pecos Texas. Southwestern Naturalist, 2, 105-121.
Mousseau, T. A. & Fox, C. W. (1998). Maternal Effects as Adaptations. Oxford: Oxford University Press.
Nakayama, S. & Miyatake, T. (2010). Genetic trade-off between abilities to avoid attack and to mate: a cost of tonic immobility. Biology Letters, 6, 18-20.
O’Steen, S., Cullum, A. J. & Bennett, A. F. (2002). Rapid evolution of escape ability in Trinidadian guppies (Poecilia reticulata). Evolution, 56, 776-784.
Partecke, J. & Schwabl, H. (2008). Organizational effects of maternal testosterone on reproductive behavior of adult house sparrows. Developmental Neurobiology, 68, 1538-1548.
Paulissen, M. A. (1998). Laboratory study of escape tactics of parthenogenetic and gonochoristic Cnemidophorus from southern Texas. Copeia, 240-243.
Pinheiro, C. E. G. (1996). Palatability and escaping ability in neotropical butterflies: Tests with wild kingbirds (Tyrannus melancholicus, Tyrannidae). Biological Journal of the Linnean Society, 59, 351-365.
Placyk, J. S. (2012). The role of innate and environmental influences in shaping antipredator behavior of mainland and insular gartersnakes (Thamnophis sirtalis). Journal of Ethology, 30, 101-108.
Price, A. H. (1992). Comparative behavior in lizards of the genus Cnemidophorus (Teiidae), with comments on the evolution of parthenogenesis in reptiles. Copeia, 323-331.
Punzo, F. (2007). Sprint speed and degree of wariness in two populations of whiptail lizards (Aspidoscelis tesselata) (Squamata Teiidae). Ethology Ecology & Evolution, 19, 159-169.
Riechert, S. E. & Hedrick, A. V. (1990). Levels of predation and genetically based antipredator behavior in the spider, Agelenopsis aperta. Animal Behaviour, 40, 679-687.
Robert, K. A., Vleck, C. & Bronikowski, A. M. (2009). The effects of maternal corticosterone levels on offspring behavior in fast- and slow-growth garter snakes (Thamnophis elegans). Hormones & Behavior, 55, 24-32.
Ruuskanen, S. & Laaksonen, T. (2010). Yolk hormones have sex-specific long-term effects on behavior in the pied flycatcher (Ficedula hypoleuca). Hormones and Behavior, 57, 119-127.
Sasaki, K., Fox, S. F. & Duvall, D. (2009). Rapid evolution in the wild: Changes in body size, life-history traits, and behavior in hunted populations of the Japanese mamushi snake. Conservation Biology, 23, 93-102.
Schall, J. J. & Pianka, E. R. (1980). Evolution of escape behavior diversity. American Naturalist, 115, 551-566.
Shaffer, L. R. & Formanowicz, D. R. (2000). Sprint speeds of juvenile scorpions: Among family differences and parent offspring correlations. Journal of Insect Behavior, 13, 45-54.
Shine, R. (1995). A new hypothesis for the evolution of viviparity in reptiles. American Naturalist, 145, 809-823.
Shine, R. & Downes, S. J. (1999). Can pregnant lizards adjust their offspring phenotypes to environmental conditions? Oecologia, 119, 1-8.
Shine, R. & Harlow, P. S. (1996). Maternal manipulation of offspring phenotypes via nest-site selection in an oviparous lizard. Ecology, 77, 1808-1817.
Shine, R., Madsen, T. R. L., Elphick, M. J. & Harlow, P. S. (1997). The influence of nest temperatures and maternal brooding on hatchling phenotypes in water pythons. Ecology, 78, 1713-1721.
Sies, H. (1997). Oxidative stress: Oxidants and antioxidants. Experimental Physiology, 82, 291-295.
Sinervo, B. & Huey, R. B. (1990). Allometric engineering: An experimental test of the causes of interpopulational differences in performance. Science, 248, 1106-1109.
Sinervo, B. & Lively, C. M. (1996). The rock-paper-scissors game and the evolution of alternative male strategies. Nature, 380, 240-243.
Sinervo, B., Svensson, E. & Comendant, T. (2000). Density cycles and an offspring quantity and quality game driven by natural selection. Nature, 406, 985-988.
Sinn, D. L., Apiolaza, L. A. & Moltschaniwskyj, N. A.(2006). Heritability and fitness-related consequences of squid personality traits. Journal of Evolutionary Biology, 19, 1437-1447.
Sorci, G., Swallow, J. G., Garland, T. & Clobert, J. (1995). Quantitative genetics of locomotor speed and endurnace in the lizard Lacerta vivipara. Physiological Zoology, 68, 698-720.
Stapley, J. & Keogh, J. S. (2005). Behavioral syndromes influence mating systems: Floater pairs of a lizard have heavier offspring. Behavioral Ecology, 16, 514-520.
Storfer, A. & Sih, A. (1998). Gene flow and ineffective antipredator behavior in a stream-breeding salamander. Evolution, 52, 558-565.
Storm, J. J. & Lima, S. L. (2010). Mothers forewarn offspring about predators: A Transgenerational maternal effect on behavior. American Naturalist, 175, 382-390.
Tobler, M., Healey, M. & Olsson, M. (2011). Digit ratio, color polymorphism and egg testosterone in the Australian painted dragon. Plos One, 6, 7.
Tobler, M., Healey, M. & Olsson, M. (2012). Digit ratio, polychromatism and associations with endurance and antipredator behaviour in male painted dragon lizards. Animal Behaviour, 84, 1261-1269.
Tocher, D. R. (2003). Metabolism and functions of lipids and fatty acids in teleost fish. Reviews in Fisheries Science, 11, 107-184.
Uller, T. & Olsson, M. (2006). Direct exposure to corticosterone during embryonic development influences behaviour in an ovoviviparous lizard. Ethology, 112, 390-397.
Wahle, R. A. (1992). Body size dependent antipredator mechanisms of the American lobster. Oikos, 65, 52-60.
Watkins, T. B. & McPeek, M. A. (2006). Growth and predation risk in green frog tadpoles (Rana clamitans): A quantitative genetic analysis. Copeia, 2006, 478-488.
Webb, J. K., Brown, G. P. & Shine, R. (2001). Body size, locomotor speed and antipredator behaviour in a tropical snake (Tropidonophis mairii, Colubridae): The influence of incubation environments and genetic factors. Functional Ecology, 15, 561-568.