ESCAPING FROM PREDATORS An Integrative View of Escape Decisions (2015)
Part III Related behaviors and other factors influencing escape
11 Determinants of lizard escape performance: decision, motivation, ability, and opportunity
Kathleen L. Foster, Clint E. Collins, Timothy E. Higham, and Theodore GarlandJr.
11.1 Introduction
All animal behavior involves movement, and most behavior involves locomotion, i.e., the act of self-propulsion. Arguably, “Locomotion, movement through the environment, is the behavior that most dictates the morphology and physiology of animals” (Dickinson et al. 2000). In other words, natural and sexual selection often act on locomotor performance abilities because they are crucial to success in many behaviors, including avoiding or escaping from predators. As Damon Runyon put it, “It may be that the race is not always to the swift nor the battle to the strong, but that is the way to bet” (Runyon 1992). Or, as noted by Woakes and Foster (1991, back cover), “Exercise lies at the heart of the struggle for existence. The exercise abilities of animals are constantly being refined by the relentless process of natural selection.” Motivated by this perspective, several artificial selection experiments have targeted aspects of locomotor behavior or performance (Feder et al. 2010; Careau et al. 2013).
The types of locomotion used while escaping from predators are highly diverse because the animals themselves, both predators and prey, are diverse. Moreover, numerous ecological factors, including gross characteristics of the habitat (e.g., presence of trees, boulders, sand) and environmental characteristics (e.g., temperature, wind, shade) are highly variable. Locomotion and its relationships to such factors ultimately determine whether a potential prey individual escapes from an encounter with a predator (Figure 11.1).
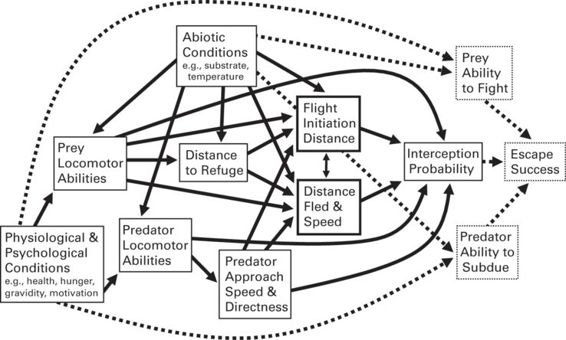
Figure 11.1
Path diagram depicting hypothesized causal relations among various factors that affect whether a prey is able to escape from a predator (see also Bulova 1994). Note that “prey ability to fight,” “predator ability to subdue,” and “escape success” are beyond the scope of this volume, and so are depicted in boxes with dotted borders and connected by dotted arrows. Various physiological and psychological conditions (bottom left) will affect the locomotor abilities of both prey and predator, and these abilities are also affected by abiotic conditions, especially in ectothermic poikilotherms. Prey locomotor abilities affect decisions about how far they stray from safe refuge and how closely they allow a detected predator to approach (i.e., flight initiation distance [FID]). Both FID, distance fled (DF), and speed affect the probability that the predator intercepts the prey, as do the locomotor abilities of both predator and prey. Once contact is made, the relative abilities of predator and prey to subdue or fight (or possibly startle or bluff) determine whether the prey is able to escape. Physical performance abilities during the “tooth and claw” phase are also affected by physiological and psychological conditions. A number of other likely effects are depicted but not discussed in this legend (see text). In addition, several important factors are not depicted, including predator crypsis and prey ability to detect predators, and prey crypsis and predator ability to detect prey, all of which are affected by both characteristics intrinsic to the organisms (e.g., coloration, visual acuity) and external conditions (e.g., wind, cloud cover, time of day; Bulova 1994). Note also that predator detection abilities, locomotor abilities, and abilities to subdue could all affect how and when the prey reacts if the prey assess this information prior to making the decision. Finally, absolute or relative body size of predator and prey can have pervasive effects on many elements within this network.
Lizards have been the subject of many studies of both locomotion and antipredator behavior. They provide a rich source for studying the ecology, performance, and biomechanics of escaping, given the disparate ecomorphological adaptations that characterize many lizard groups (Pianka & Vitt 2003). For example, geckos and anoles have adhesive toepads that allow them to escape on vertical and even inverted surfaces (Russell 1975). Chameleons have prehensile feet that allow them to grip thin branches during escape (Higham & Anderson 2013). In sandy desert habitats, several lizard species have convergently evolved toe fringes and/or webbed feet that increase surface area (Carothers 1986; Luke 1986; Bauer & Russell 1991). Basilisks have evolved similar foot structures to allow them to run across water rather than sand. Flying lizards can glide using foldable “wings” constructed of skin stretched across their ribs. This remarkable phenotypic variation should be considered when determining how, where, and when lizards might escape.
Lizards are diverse and flee by various means, including running, jumping, climbing, gliding (McGuire 2003), swimming, and diving. Aside from fleeing, lizards have alternative ways to avoid or cope with predators, such as crypsis, armor, fighting, blood-squirting, and venom (e.g., John-Alder et al. 1986; Sherbrooke & Middendorf 2001). As our experience is mainly with lizards, we focus on them in this chapter although most of the concepts that impact locomotor performance and escape success are more broadly applicable to other terrestrial vertebrates. We will briefly discuss how such concepts as detection of predators, decisions that lizards must make, and flight initiation distance relate to and are determined by locomotor performance (Figure 11.1), but our main focus is the morphological and physiological basis of performance limits in the context of escape success.

Figure 11.2
Lizards exhibit a great diversity of escape behaviors, including burrowing, sand-swimming, jumping, running, swimming, diving, escaping into crevices, climbing, and even “flying.” Shown here are (clockwise from top left) a fringe-toed lizard (Uma sp.), collared lizard (Crotaphytus collaris), tokay gecko (Gekko gecko; photo credit: Lee Grismer), venomous Gila monster (Heloderma suspectum; see John-Alder et al. 1986), horned lizard after squirting blood from its orbital sinus (Phrynosoma sp.), sand lizard shimmying into sand (Meroles sp; photo credit: Clint E. Collins), tree lizard (Lacerta viridis; photo credit: Robert & Mihaela Vicol), flying lizard in the middle of a glide (Draco taeniopterus; see McGuire 2003), and chuckwalla near rock crevices into which it retreats when threatened (Sauromalus sp; photo credit: Clint E. Collins). Unless otherwise specified, images were obtained from Wikipedia Commons.
11.2 Locomotor performance affects escape decisions
11.2.1 Temperature effects on fight or flight
Once a predator is detected, the prey has to make a decision about whether it will flee or rely on some other mechanism for defense. Temperature is a primary factor that appears to determine whether a lizard will fight or flee because locomotor performance is (up to a point) increased at warmer body temperatures (e.g., sprint speed: Herrel et al. 2007; endurance: Garland 1994). Two species of agamid lizards living in open habitats where refuges are scarce attempt to flee when warm, but stand and fight when temperatures are cooler and sprint speed is too slow to permit escape (Hertz et al. 1982; see also Crowley & Pietruszka 1983). The physiological basis for this temperature-dependent shift in antipredator behavior appears to center on the differences in temperature dependence of force production in the locomotor and biting muscles of these lizards (Herrel et al. 2007). Although locomotor muscles generate peak force under a relatively narrow, warmer range of temperatures, the adductor mandibulae externus superficialis posterior, one of the jaw muscles responsible for producing force during biting, is largely temperature independent, generating near maximal forces over a wide range of temperatures (Herrel et al. 2007).
11.2.2 Impacts of locomotor performance on flight initiation distance
Fleeing from predation has largely been understood through the framework of models utilizing economic (cost-benefit) escape theory (Ydenberg & Dill 1986; Cooper & Frederick 2007, Chapter 2). As described in Chapters 1 and 2, an animal detects a predator and then monitors the predator as it approaches, fleeing when the cost of remaining in place equals the cost of fleeing (Ydenberg & Dill 1986) or the expected post-encounter Darwinian fitness is maximized (Cooper & Frederick 2007). The distance between the predator and the prey when the animal initiates its escape is termed the flight initiation distance (FID). Evading predation via locomotion involves complex, high-power movements, including jumping, accelerating, running, and evasive maneuvers (Djawdan 1993; Zehr & Sale 1994; McElroy et al. 2007; Higham & Irschick 2013). Therefore, evading predators requires high power output from skeletal muscles while operating within the biomechanical constraints that limit performance. Biomechanical and physiological sources of variation in performance often affect FID in ways consistent with escape theory. For example, individual lizards with cooler body temperatures escape more slowly relative to warmer counterparts (Cooper 2000; Cooper & Frederick 2007; Herrel et al. 2007). To compensate for decreased locomotor capacity, cooler lizards increase FID, presumably to prevent their predators from overtaking them (Rand 1964; Chapter 5). Underlying increased FID and reduced locomotor capacity at cooler temperatures is the sensitivity of muscle function to temperature, reduced metabolic rate, and reduced enzymatic activity rate (Adams 1985; Marsh & Bennett 1986a, b; Rome and Bennett 1990; Fitts et al. 1991; Dickinson et al. 2000; Jayne & Daggy 2000). For example, the frequency of muscle contraction at which mechanical output is greatest, known as optimal cycling frequency, increases with temperature. However, increasing temperature beyond a certain range may ultimately reduce power output (reviewed in James 2013) and hence locomotor abilities. Thus the interaction between temperature and locomotor physiology can impact not only escape performance, but also decisions about when to initiate escape behaviors.
Variable habitats often contain physical structures that constrain escape behavior and performance, and likely necessitate changes in locomotor kinematics, which could play an important role in determining FID. For example, increases in substrate rugosity or compliance likely constrain a lizard’s ability to achieve maximal sprint speeds during escape, thus potentially increasing its FID or requiring an altered trajectory to minimize encounters with substrates that would reduce performance (Figure 11.3; Irschick & Losos 1999; Tulli et al. 2012; Collins et al. 2013). Given that terrestrial and arboreal environments present a diversity of obstacles, lizards that use highly complex habitats likely have evolved mechanisms for achieving high performance despite structural variability. For example, lizards, including Anolis carolinensis, Callisaurus draconoides, Uma scoparia, and some species of Varanus, alter limb posture and/or kinematics to deal with decreases in perch diameter and/or increases in perch incline, and these kinematic changes affect downstream changes in locomotor speed (Irschick & Jayne 1999; Jayne & Irschick 1999; Higham & Jayne 2004a, b; Spezzano & Jayne 2004; Foster & Higham 2012; Clemente 2013). Kinematics also play an important role in enhancing or constraining locomotor performance during escape because postural shifts may alter the ability of elastic tendons to store energy and/or may dictate the operating lengths of muscles (Higham & Irschick 2013). For example, different species of Anolis lizards have different levels of elastic energy storage prior to jumps because their femoral protraction differs (Vanhooydonck et al. 2006b, c). Further, whereas Anolis lizards decrease limb cycling rate to increase stability on narrow perches (Foster & Higham 2012), Varanus lizards achieve both high sprint speeds and stability by increasing stride frequency (Marsh & Bennett 1986a, b; Clemente 2013). This highlights the idea that there are likely multiple, context-dependent solutions to predator evasion (Wainwright et al.2005; Wainwright 2007; Bergmann et al. 2009; Garland et al. 2011). Hence, it is likely that a strong relationship exists between FID and the ability to make kinematic and behavioral adjustments to compensate sufficiently for reduced performance.
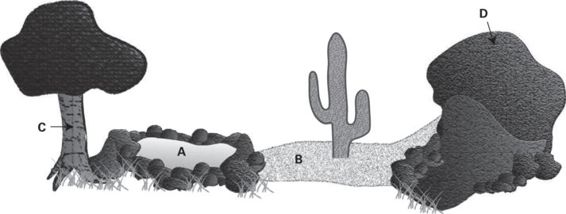
Figure 11.3
Schematic showing the variety of substrate textures or rugosities commonly encountered by lizards. Substrates can be very smooth and/or compliant (e.g., water [A], and sand [B]) or firm with varying degrees of roughness (e.g., tree bark [C] and rock [D]). Many lizards have pedal specializations that assist locomotion on these substrates: geckos have adhesive toe pads that are very effective at clinging to extremely smooth surfaces such as leaves and windows (although they cannot stick to Teflon), sand-dwelling lizards or juvenile basilisks may have toe-fringes or webbed feet to increase surface area for moving on fluid surfaces, and claws help lizards move on rough surfaces.
Changes in performance capability may underlie ontogenetic differences in FID in lizards. Smaller, younger individuals are slower (Garland 1985), and thus may rely on either longer FID or crypsis to survive. Conversely, adult lizards can increase FID to save the energy that would be expended on a longer, faster run. Nevertheless, the patterns of FID, crypsis, and performance capabilities are nuanced and context specific; for example, in the absence of protective refuges, juveniles flee more frequently than adults (Smith 1997). Future studies should determine how selection acts on the integrated suites of acceleration, agility, sprint speed, and behavior to determine the true modus operandi of selection on locomotion. Finally, to compensate for hypothetically lower performance capabilities, juvenile lizards may use a greater percentage of their maximal sprinting and accelerating capabilities during escape than their adult counterparts (Irschick 2000; Toro et al. 2003; Stiller & McBrayer 2013). This variation suggests motivational differences may be key.
11.3 Motivation: how do animals decide what to do?
Motivation can strongly affect the performance exhibited by an individual in a given locomotor event (Astley et al. 2013). Performance is defined as the “ability of an individual to conduct a task when maximally motivated” (Careau & Garland 2012, p. 546). We define the motivation of an animal as the sum of factors or conditions that (1) stimulate or arouse an animal to perform a given task and (2) determine the level of persistence and vigor as it performs that task (modified from Beck 1978). Motivation in lizards has been inferred from measures of speed, latency between stimulus and response, and the number of stimuli required to elicit a response (e.g., Sorci et al. 1995; Skelton et al.1996). However, these indirect measures may not allow identification of the factor or stimulus that is most important for motivating a lizard to run. Attempts to assess motivation in the field under a variety of natural and ecologically relevant conditions would help identify the situations and motivating factors associated with high performance.
Once a predator has been detected, the motivation of the prey is an integral factor in deciding whether or not to flee and in the subsequent level of locomotor performance during the escape attempt. Interpopulational differences in sprint capacity and/or voluntary running speed have been found within species of Podarcis, Sceloporus, and Tropidurus: individuals in populations exposed to greater predation risk may have greater sprint speeds (Crowley 1985a, b; Snell et al. 1988; Van Damme et al. 1998; but see Huey & Dunham 1987; Huey et al. 1990). However, many species of lizards do not use their maximum performance capacity when escaping from a predator (Hertz et al. 1988; Garland & Adolph 1991; Irschick & Losos 1998; Irschick & Garland 2001; Irschick 2003; Irschick et al. 2005; Husak 2006b; Husak & Fox 2006). This may be a function of motivational differences and may help to explain differences between maximum locomotor performance measured in the laboratory and realized field performance (Bennett & Huey 1990; Huey et al. 1990; Irschick 2003; Irschick et al. 2005). Thus, in some cases, lizards may be more motivated to run away from a human “predator” in a laboratory setting than they are to run from a common natural predator in the wild.
To be successful, prey must be able to effectively forage and attract mates. The conflicting demands imposed by multitasking may contribute to a prey’s level of motivation when fleeing. Lizards may be reluctant to escape if their physiological state (e.g., gravidity, injury, tail loss, body size, time since last meal) or the abiotic conditions (e.g., habitat structure, temperature, humidity) are such that their locomotor performance capacity is diminished. Beyond these considerations, personality traits that dispose an animal to take greater or lesser risks may contribute to motivation regarding types and levels of performance during escape (Cooper 2008; Careau & Garland 2012). Unfortunately, intuitive, concrete experimental data supporting the impact of motivation on escape performance in ecologically relevant contexts are scarce.
11.4 Escape ability
The potential level of performance a lizard reaches in a given escape attempt is limited by its ability at that time, which is affected by various morphological and physiological traits (Figure 11.4). All aspects of locomotor ability are likely to be interrelated to some extent because each subcategory of ability is influenced by such factors as body size, limb lengths and proportions, muscle size and composition, innervation, neural control, and tendon characteristics (Garland 1993; Garland & Losos 1994; Christian & Garland 1996; Higham & Irschick 2013; Figure 11.4). Many of these relationships may cause trade-offs or constraints (Garland 2014) on the development or evolution of locomotor abilities (Vanhooydonck et al. 2014). Another source of constraint is the conflicting functions of animal phenotypes (Higham & Irschick 2013). This is the case for breathing and running in some lizards, where hypaxial and epaxial muscles drive both the movements of the ribs (for breathing) and of the body (for locomotion) (Carrier 1991; Farmer & Carrier 2000a, b; Brainerd & Owerkowicz 2006). The dual role of these muscles would appear to be one important factor that can limit the endurance capacity of a lizard, and therefore its escape ability in some situations.
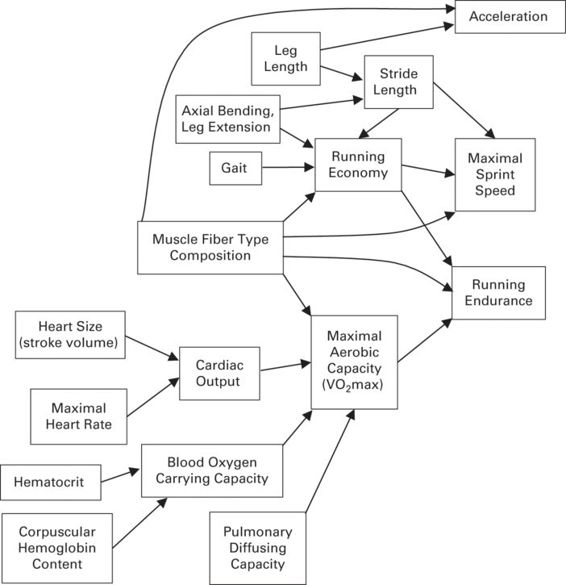
Figure 11.4
Path diagram illustrating interrelationships among some of the phenotypic traits that affect locomotor performance abilities, three of which are depicted (acceleration, sprint speed, and running endurance).
11.4.1 Body size and shape
How the evolution of morphological diversity relates to variation in animal performance is a fundamental question in both functional and evolutionary biology. This section will contextualize lizard morphological diversity in relation to escape diversity and attempt to link form with function. An overarching perspective is that body size has pervasive influences because it (1) affects the mass an animal must move and (2) is often allometrically associated with form (e.g., leg length: Christian and Garland 1996; but see McGuire 2003), thus having indirect effects on anything that form affects. Therefore body size strongly impacts lizard locomotor performance and escape behavior (Figure 11.5). Moreover, body size itself often shows complicated evolutionary patterns even within relatively uniform clades of squamates (e.g., McGuire 2003; Collar et al. 2011).
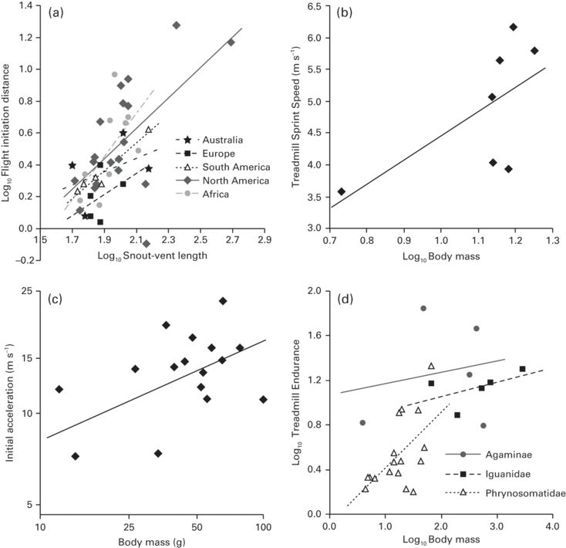
Figure 11.5
Relationships between body size and several traits relevant to escape performance. (A) Snout-vent length vs. flight initiation distance across several lizard taxa from continental Australia, Europe, South America, North America, and Africa (data from Cooper et al. 2014). (B) Body mass vs. sprint speed (measured on a treadmill) among seven Cnemidophorus species (data from Bonine & Garland 1999). (C) Body mass vs. acceleration (at 0.05 s) in the agamid lizard Stellio stellio (redrawn from Huey & Hertz 1984). (D) Body mass vs. endurance (measured on a treadmill) in three groups of iguanian lizards (data from Garland 1994).
Hatchling and juvenile lizards are generally thought to be under relatively strong selection owing to their small size and limited locomotor capacity (Wassersug & Sperry 1977; Carrier 1996; Husak 2006a). However, one advantage smaller, younger lizards may have over adults is the ability to utilize a wider range of habitat structures and to escape into smaller retreats. For example, unlike adults, juvenile basilisks (Basiliscus basiliscus) are able to escape by running over water surfaces because the smaller juvenile lizards can generate greater force relative to their body size (Glasheen & McMahon 1996). Similarly, smaller Anolis lizard individuals utilize a wider range of habitat structures than their larger counterparts (Irschick & Losos 1999). How juveniles utilize acceleration and agility throughout ontogeny may be of utmost importance, especially given the differential impact of slopes and habitat structure on size. Although larger lizards can sprint and accelerate faster than smaller counterparts (Huey & Hertz 1984), smaller lizards may have smaller turning radii and thus outmaneuver larger predators, a prediction that deserves empirical testing.
Regardless of the cause of size differences (ontogeny, phylogeny, sexual dimorphism, etc.), steep inclines significantly reduce sprint performance in large animals relative to smaller animals (Jayne & Irschick 2000; Birn-Jeffery & Higham 2014). For example, the striped plateau lizard (Sceloporus virgatus) flees shorter distances upslope than horizontally or downslope (Cooper & Wilson 2007). If a larger animal were to chase the lizard upslope, it would have to expend more energy and would be at a disadvantage due to the greater work required to move its larger body uphill, as has been shown in lizards, in both the laboratory and the field (Huey & Hertz 1984; Jayne & Irschick 2000; Birn-Jeffery & Higham 2014).
Sexual differences in escape ability (e.g., sprint performance) are often attributed to the energetic costs of gravidity (Shine 2003a, b). For example, increased body mass alone could cause gravid females to have reduced locomotor abilities (Garland 1985; Garland & Else 1987). However, because reduced locomotor performance likely decreases future opportunities for reproduction, compensating for gravidity-induced impairment would have obvious evolutionary significance (Arnold 1983; Reznick 1985; Brodie 1989; Husak, 2006a, b). When performance and escape differences cannot be attributed to size alone, lower-level physiological differences are often the answer (Shine 2003a, b; Lailvaux 2007; Husak & Fox 2008). Females that are less conspicuous than males can rely relatively more on crypsis compared to males to compensate for lower sprinting capacities (Cooper et al. 1990; Cooper & Vitt 1991). In fact, whereas non-gravid female collared lizards (Crotaphytus collaris) compensate for reduced locomotor performance by using a greater proportion their lab-tested sprint speed, gravid collared lizards remain closer to refuges relative to non-gravid counterparts. Further, when they do escape, females may use a greater proportion of their maximum sprint capacity to achieve similar escape speeds in the field (Husak 2006b, c; Husak & Fox 2008). Interestingly, although many studies have supported the hypothesis that gravidity temporarily constrains escape capacity due to increased mass (Le Galliard et al. 2003), gravid green iguanas (Iguana iguana) increase mechanical power, peak forces, step duration, and limb swing speed to compensate for the greater mass (Scales & Butler 2007). Furthermore, differences in escape ability and decisions may be attributed to conflicts between optimal temperatures for eggs and optimal escape temperatures (Lailvaux et al. 2003). Finally, Irschick et al. (2003) found that at least one gecko, Hemidactylus garnotii, actually ran faster with an added load equaling 2% of its body weight (although sprint speed decreased with further increases in load). This non-linear effect suggests that there may be an optimal loading weight that may be related to gravidity. Future studies could compare optimal vs. actual reproductive mass increases.
The evolution of body shape is another important factor in determining escape response and success. In many animals, stockiness constrains flexibility and thus limits the contribution of axial movements to locomotion (Brainerd & Patek 1998; Walker 2000; Bergmann et al. 2009). Because many tetrapods, including lizards, rely on axial bending to increase stride length, and thereby sprint speed and endurance, reduced flexibility of the axial skeleton hypothetically decreases locomotor performance (Reilly & Delancey 1997; Reilly 1998; Walker 2000). Therefore, to evade predators, lizards with relatively stocky bodies should exhibit morphological and behavioral compensations for this locomotor impairment. One clear example is the evolution of stockiness in the genus Phrynosoma, which compensates for reduced locomotor capabilities with greater morphological investment in armor (horns) and crypsis, in addition to the unique ability to squirt blood from the orbital sinus (Sherbrooke & Middendorf 2001). Conversely, cursorial sand lizards, such as the zebra-tailed lizard (Callisaurus draconoides), have evolved slender bodies with longer legs and tails. Lizards at this end of the spectrum rely on high speeds to evade predators and do not possess armor as Phrynosoma does (Figure 11.6A). Furthermore, the medial iliofibularis, a swing phase locomotor muscle, consists primarily of fast-twitch oxidative glycolytic fibers in Phrynosoma and fast-twitch glycolytic fibers in the cursorial sand lizards (Bonine & Garland 1999; Bonine et al. 2001, 2005). This trade-off indicates that stockier, armored lizards such as Phrynosoma rely more on slower, aerobically powered steady locomotion that is unrelated to predation pressure, rather than high-powered, anaerobic bursts (Figure 11.6B).
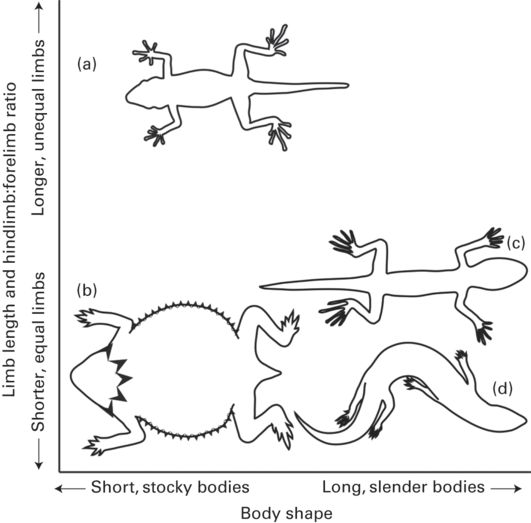
Figure 11.6
Schematic illustrating the relationship between hindlimb to forelimb ratio and body shape among lizards. (A) Rhoptropus afer is specialized for higher running speeds and has longer hindlimbs relative to forelimbs. (B) Phrynosoma cornutum exemplifies the short-limbed, stocky, and heavily armored phenotype that has reduced mobility. (C) Arboreal Cnemaspis species, like many arboreal lizards, tend to have more equal forelimb and hindlimb lengths, reflecting the increased propulsive importance of the forelimbs when climbing, and slender bodies for greater stability on narrower branches. (D) Scincus scincus has a hydrodynamic, slender body and short, relatively equal forelimbs and hindlimbs to facilitate sand-swimming.
In constrast with the sand lizards, slender-bodied skinks and alligator lizards have reduced limb lengths (John-Alder et al. 1986; Bonine & Garland 1999). Shorter legs and elongate bodies may allow easier navigation through leaf litter, in-ground burrows, and tall grasses. Escape into such microhabitats may confuse predators or prevent them from following (Jaksić & Núñez 1979; Melville & Swain 2000; Melville 2008). Although it is thought this escape behavior enhances the probability of escape (Schall & Pianka 1980), little is known about other axial or appendicular adaptations for increased turning and maneuverability in lizards. Although turning generally decreases sprint speed in some lizards (Jayne & Ellis 1998; Irschick & Jayne 1999; but see Higham et al. 2001), it is likely that behavioral and performance responses to turning (Howland 1974) vary by species and habitat type. However, this remains to be explored.
Another body shape modification for escaping in habitats that impose intense functional constraints is body flattening by some saxicolous lizards (Goodman 2007, 2009; Revell et al. 2007; Goodman et al. 2008). Although some saxicolous species exhibit morphological specializations for highly rugose cliffs and skree, high-speed locomotion on vertical rock faces is difficult, so many saxicolous species hide within narrow crevices to avoid or escape from predators (Revell et al. 2007; Goodman et al. 2008; Tulli et al. 2012; Collins et al. 2013).
11.4.2 Limb and muscle morphology
Limb length strongly impacts locomotor performance; lizards with longer hindlimbs and shorter forelimbs achieve greater sprint speeds and jump farther (e.g., Bonine & Garland 1999; Toro et al. 2004) although there may be additional advantages to longer limbs that are unrelated to locomotor performance (Iraeta et al. 2011). However, longer limbs may not always be optimal for effective escape from a predator. As discussed above, shorter limbs may be advantageous for maneuvering in confined spaces, such as burrows, because they may achieve faster cycling frequencies and are easier to maneuver around obstructions. Shorter limbs may also be beneficial for stability on narrower surfaces. In Anolis, although shorter limbed species don’t run faster on narrower surfaces than longer limbed species (Irschick & Losos 1999), they may be more effective at escaping if shorter limbs reduce the tendency to fall. This dual requirement of speed and surefootedness in order to achieve effective escape in arboreal situations is illustrated in Sceloporus occidentalis, in which terrestrial populations with longer limbs run slower than shorter limbed arboreal populations on narrow substrates (Figure 11.7; Sinervo & Losos 1991).
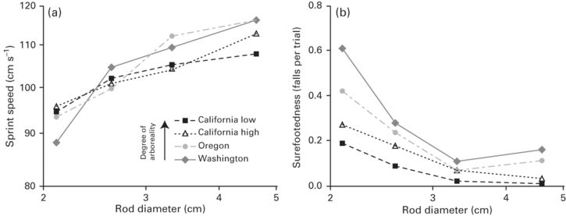
Figure 11.7
Impact of substrate diameter on (A) sprint speed and (B) surefootedness of four populations (California [high and low elevation], Oregon, and Washington) of the western fence lizard, Sceloporus occidentalis, that are arboreal to varying degrees. The more arboreal populations sprint faster and are more stable on narrowest perches compared to the more terrestrial populations.
(Redrawn from Sinervo & Losos 1991)
Differential elongation of limb segments may also affect locomotor performance. Elongation of distal limb segments relative to proximal segments is an indication of cursoriality (Coombs 1978; Hildebrand 1985) and the metatarsal:femur ratio is correlated with maximal sprint speed in mammals (Garland & Janis 1993). However, the effect of different relative limb segment lengths on lizard locomotor performance remains unclear. Van Damme et al. (1998) found a slightly longer foot length in faster species, but this difference was unlikely to have caused the two-fold increase in speed among the species examined (Aerts et al. 2000). Similarly, contrary to expectations, limb segment length, independent of total limb length, is not correlated with jumping performance in Anolis (Toro et al. 2004). In bipedal lizards, the story gets a little more interesting. In addition to longer tails, shorter forelimbs (especially the manus), and longer hindlimbs (except in some agamids), bipedal lizards either have disproportionately longer proximal hindlimb segments (in iguanids) or distal (pes) hindlimb segments (Snyder 1954, 1962). However, although lizards that can run bipedally generally run faster than strictly quadrupedal lizards, these morphological differences may be a function of the different mechanics of the two types of gaits. Based on what we know about mammalian limb morphology and segment ratios, we can hypothesize certain morphologies in lizards specialized for high sprint speeds vs. those with limbs specialized for greater strength (Figure 11.8A). However, more research into the impact of limb morphology and segment length on lizard locomotor performance is necessary to test these hypotheses.
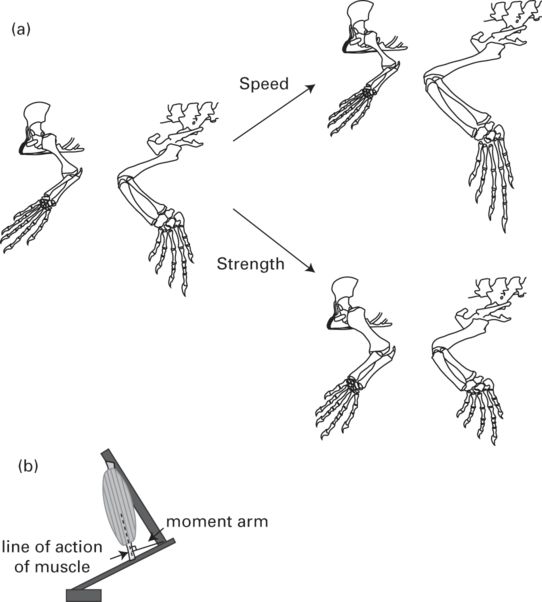
Figure 11.8
(A) Left: forelimb and hindlimb of Varanus sp., after Bellairs (1970). Relative size of forelimb and hindlimb approximately scaled to reflect average ratio of forelimb:hindlimb segment length ratio of 22 varanid species from Christian and Garland (1996). Top right: hypothesized morphology of lizard with fast sprint speed. Note elongation of hindlimb segments, especially phalanges, and shortening of forelimb segments, especially manus. Bottom right: hypothesized morphology of lizard specialized for strength, as might be important during digging. Note shortening and thickening of limb segments, lengthening of forelimb, and shortening of hindlimb. (B) Schematic showing calculation of muscle moment arm. The moment arm is the perpendicular distance between the line of action of the muscle and the joint at which the muscle acts.
Research into these gross external morphological features should be complemented by examination of internal muscle morphology, as muscles are the functional units responsible for powering locomotion. Muscles are complex and can vary in both morphology and the mechanics of contraction, both of which can have a profound impact on locomotor capacity (Loeb & Gans 1986; Biewener 1998; Lieber & Ward 2011). Here, we will briefly discuss how the morphological aspects of muscle may affect locomotor performance. A discussion of relevant physiological aspects of muscle will follow in section 11.4.4.
The force a muscle can generate is a function of its physiological cross-sectional area (PCSA) because with a greater PCSA, a greater number of sarcomeres in parallel can contract simultaneously to contribute to force, assuming maximal stimulation (Haxton 1944; Alexander 1977; Sacks & Roy 1982; Loeb & Gans 1986; Zaaf et al. 1999; Herzog 2000; Allen et al. 2010; Lieber & Ward 2011). The length of muscle fibers affects total mechanical power because with longer fibers, more sarcomeres can be arranged in series and greater changes in length (and therefore increases in speed) are possible (Loeb & Gans 1986; Gans & de Vree 1987; Biewener 1998; Biewener & Roberts 2000; Allen et al. 2010; Lieber & Ward 2011). The placement of the muscle, i.e., the origin and insertion points of the muscle relative to the joints across which the muscle acts, also can affect locomotor function. When the distance between the joint and the line of action of the muscle is greater, the muscle has a greater moment arm, and this increases the mechanical advantage of the muscle such that less muscular force is required to generate movement about the joint (Figure 11.8B; Gans & de Vree 1987; Richmond 1998; Rassier et al. 1999; Zaaf et al. 1999; Payne et al. 2006; Wilson & Lichtwark 2011). However, increasing the muscle’s moment arm has a negative impact on the angular displacement and velocity that can be achieved about the joint because a given change in muscle length generates a smaller change in joint angle (Rassier et al. 1999; Payne et al. 2006; Wilson & Lichtwark 2011). These relationships lead to a trade-off between force and velocity such that larger muscle moment arms result in greater force, but lower velocity. Thus we would expect to see smaller moment arms in limbs that need to achieve high angular velocities, as might be beneficial for increasing stride frequency, but larger moment arms in limbs that are built for strength, as might be beneficial in climbing and burrowing species. Although these predictions may hold in small-scale comparisons (Zaaf et al. 1999; Herrel et al. 2008), broad comparative studies of muscle morphology (in the context of maximum performance) are lacking for lizards. Do lizards that sprint faster (and cycle their limbs faster) exhibit decreased moment arms compared to those lizards that move slower when escaping? Such studies will likely be particularly important because they have the potential to incorporate the ecologically relevant comparison of prey vs. predator morphology and performance.
11.4.3 Escape performance and tail autotomy
Many lizards possess the ability to autotomize (sever) the tail during a predator-prey interaction. The lost tail can provide a meal for the predator. If neither the tail nor the body are captured (Bellairs & Bryant 1985), the autotomized tail may move dramatically, distracting the predator while the lizard flees (Higham & Russell 2010, 2012; Higham et al. 2013a, b). Caudal autotomy often enables lizards to survive a predatory attack (e.g., Daniels et al. 1986). In some circumstances, the autotomized tail itself evades the predator completely, allowing the lizard to return to the site of autotomy to ingest the lost tail, which contains fat reserves (Clark 1971). Below, we discuss the impact of caudal autotomy on locomotor behaviors critical to escape success: running and jumping.
Although the tail of lizards is critical for locomotor movements, such as running and jumping, losing a tail is beneficial enough to offset the costs associated with modifications of locomotor movements. Running fast is an obvious mechanism for evading a predator, and the tail plays a significant role in locomotor mechanics of terrestrial lizards. A question that has arisen multiple times is whether running speed increases or decreases following autotomy. Skinks and iguanids exhibit a consistent decrease in running speed following autotomy, whereas geckos and lacertids are highly variable, with some geckos exhibiting a drastic increase (Daniels 1983; McElroy & Bergmann 2013). Interestingly, lizards with larger tails tend to exhibit a greater decrease in speed (McElroy & Bergmann 2013).
Jumping is important during escape for many lizards, especially those in arboreal habitats (Losos & Irschick 1996; Higham et al. 2001), and tails can be critical for maintaining in-air stability (Gillis et al. 2009, 2013). Anolis lizards with intact original tails took off and landed with approximately the same body angle. In contrast, tailless anoles underwent significant posterior rotation, up to 90° between take-off and landing (Gillis et al. 2009). Further, although lizards often autotomize only a portion of the tail, there was no significant impact of the amount of tail lost on body rotation during jumping (Gillis et al. 2013). However, high variation within groups that had autotomized different amounts of the tail may have obscured any effects of the proportion of the tail lost. Lizards that have lost their tail exhibit behavioral shifts to compensate for decreased stability. For anoles, this might mean occupying areas closer to the ground to avoid injury while jumping during an escape or avoiding jumping altogether. Whether such changes result in increased mortality and decreased fitness remains to be tested.
In most cases, the autotomized tail is not lost forever. The ability to regenerate the lost appendage is common, but it is not clear how the ability to escape is impacted by having a regenerated tail. The composition of the regenerated tail is quite different from the original, which suggests that function may be altered (Gilbert et al. 2013; Russell et al. 2014). It is likely that fine control of tail movements is compromised in the regenerated tail, leading to decreased stabilizing ability during jumping and/or running. Future work should examine how the function of a regenerated tail during locomotion compares with that of the original tail, but also how performance changes immediately after autotomy and during the phase of regeneration. Given that regeneration can take months, locomotor performance may gradually recover over this period of time. However, it is also possible that performance does not recover at all.
11.4.4 How muscle physiology is related to escape ability
Given that muscles actuate the movements of the limbs during escape, it is expected that muscle physiology correlates with escape performance. Despite this, few studies have linked muscle physiology with running speed in lizards (Gleeson & Harrison 1988; Higham et al. 2011a; Kohlsdorf & Navas 2012; Vanhooydonck et al. 2014). Additional layers of complexity include a number of physiological and morphological parameters that might be related to locomotor performance, as well as the number of muscles that could be examined. For example, one could quantify the relative proportion of a given fiber type (fast glycolytic [FG], slow oxidative [SO], and fast oxidative glycolytic [FOG]), enzymatic activity, cross-sectional area of the whole muscle, fiber cross-sectional area, mass, moment arm, and other subordinate traits. Both propulsive (stance phase) and recovery (swing phase) muscles could be examined, and each may give a different result. Finally, the ecological context of the locomotor event might impact the relationship between muscle physiology and performance. For example, the morphological/physiological predictors of performance on a level surface differ from those on a vertical surface in the Florida scrub lizard, Sceloporus woodi (Higham et al. 2011a). The diameter of FG fibers in the gastrocnemius was correlated with acceleration on a level surface, whereas the percentage of FG fibers in the gastrocnemius predicted acceleration on a vertical surface.
Sprinting fast is commonly associated with increased escape performance. This makes sense, given that faster speeds will potentially increase the distance between the prey and predator over a given period of time. Gleeson and Harrison (1988) linked muscle morphology and physiology (iliofibularis, gastrocnemius, and caudofemoralis) to sprint speed in the desert iguana, Dipsosaurus dorsalis. They found that up to 30% of the variation in sprint speed could be explained solely by variation in fiber cross-sectional area; decreased area was associated with faster speeds. In Tropidurus lizards, faster sprint speeds were associated with a higher proportion of FG fibers in the iliofibularis muscle (Kohlsdorf & Navas 2012). Although not tested directly, it appears that those phrynosomatid lizards that sprint faster also exhibit a greater proportion of FG fibers in the iliofibularis muscle (Bonine et al. 2001). For example, the horned lizards sprint slowly compared to other species of phrynosomatids, and they exhibit the lowest percentage of FG fibers (Bonine et al. 2001). Collectively, it appears that increases in the relative proportion of fast-twitch muscle, which is good for power generation, is associated with greater escape performance in lizards.
A key consideration when attempting to quantify escape ability in relation to muscle physiology is which aspect of performance to measure. Although sprint speed is a common metric, and is likely important for escaping, acceleration might be more important in some cases. Indeed, the ability to accelerate maximally may be crucial for initially increasing distance from the predator, whereas maximum sprint speed is likely important for maintaining that distance. Muscle physiology may be linked to acceleration performance, as opposed to sprint speed, given the relatively higher demand that acceleration places on the locomotor system. As noted above, this is true for Sceloporus woodi moving on level and inclined surfaces (Higham et al. 2011a). Muscle mass-specific power during acceleration in Sceloporus woodi approaches the maximum power output measured for lizard hindlimb muscles, 90 W kg−1(McElroy & McBrayer 2010). Future work should tease apart the relative importance of acceleration and maximum sprint speed in determining the outcome of predator-prey interactions in lizards. In addition to sprint speed and acceleration, endurance capacity may play a critical role in escaping from predators. This type of performance is likely enhanced by having a greater proportion of SO muscle fibers, which are ideal for powering behaviors requiring endurance. We know very little about the links between endurance and escape success in lizards, so studying this would be a logical first step.
Overall, the relationship between muscle and escape performance is incredibly complex, depending on the type of muscle measurement, the muscle being measured, the species being examined, the ecological context, and the performance variable being quantified. Future work that illuminates the key variables would propel our understanding of escape performance.
11.5 Opportunity: escaping in different habitats
Habitat structure and substrate characteristics can profoundly impact the successful negotiation of an animal through its environment. In this section we discuss the interesting challenges posed by sandy, arboreal and saxicolus habitats, and the potential consequences for escape performance.
11.5.1 Sandy habitats
Although sandy habitats are often considered relatively simple, they make physical demands that impose selection on many psammophilous lizards. Two well-studied challenges common to many deserts are substrate compliance and incline. Here, we will discuss the challenges and compensatory mechanisms related to substrate compliance, leaving discussion of the effects of incline for the following arboreal section.
Compliant surfaces, such as sandy flats and dunes, dissipate greater amounts of energy than non-compliant surfaces such as rocks. Consequently, escaping on compliant surfaces requires increased energy expenditure (Zamparo et al. 1992; Lejeune et al. 1998). One reason for this is the decreased ability to employ elastic elements as energy-saving mechanisms because the greater compliance interferes with effective loading of structures such as tendons. However, various adaptations, such as specialized external morphological features (e.g., toe fringes) and modified kinematics (e.g., foot posture and movement) enable effective, high-speed locomotion on compliant surfaces (Glasheen & McMahon 1996; Hsieh & Lauder 2004; Tulli et al. 2009; Li et al. 2012). Nonetheless, maximal speeds and/or acceleration of lizards running on sand may be reduced as compared with the same species running on a surface that provides good traction and is less compliant (Carothers 1986).
Laterally projected, elongated toe fringes increase toe surface area, and thus traction, on substrates such as sand and water (Carothers 1986; Luke 1986). The benefits of toe fringes were discovered through an experiment that demonstrated that ablating toe fringes reduced acceleration and sprint speed on sandy surfaces, but not on rigid surfaces (Carothers 1986). Interestingly, whereas some lizards have toe fringes to increase surface area, secondarily terrestrial geckos exhibit digit modifications that reduce surface area. Many arboreal and saxicolous pad-bearing geckos achieve adhesion by increasing their subdigital surface area to form microscopic setae, which bond with surfaces through van der Waals interactions (Autumn & Peattie 2002), but can become clogged with sand (Hansen & Autumn 2005). Thus secondarily terrestrial geckos have lost or reduced these adhesive structures, which likely allows them to perform better on flat, sandy surfaces (Lamb & Bauer 2006).
When lizards traverse non-solid surfaces, their hindlimb kinematics and foot use change to accommodate slipping and energy dissipation (Li et al. 2012). On solid surfaces the zebra-tailed lizard, Callisaurus draconoides, employs digitigrade foot posture and spring-mass mechanics, whereas on granular surfaces they use plantigrade foot posture and paddle-like foot rotation (Li et al. 2012). In addition to these kinematic modifications, sand-swimming lizards can employ unusual locomotor mechanisms to propel themselves beneath the sand surface. For example, high-speed x-ray cinematography of sand-swimming revealed that the sandfish (Scincus scincus) retracts its limbs to the sides of its body and propagates a wave down its body through lateral undulation to propel itself through the sand (Maladen et al. 2009). This study highlights the importance of integrating natural history, morphology, and biomechanical techniques to understand how evolution has acted upon integrated suites of locomotor traits.
In special cases, lizards, such as the plumed basilisk (Basiliscus plumifrons), can escape across water, which is also a compliant surface. Much like running across soft sand, elastic energy storage capacity is compromised when escaping across water since the foot actually breaches the water’s surface during the stance phase (Hsieh & Lauder 2004). Thus the limb acts like a piston, doing work throughout the stride, which is in contrast with typical terrestrial legged mechanics.
11.5.2 Arboreal habitats
The arboreal habitat may be one of the more challenging habitats; it is often highly heterogeneous in incline, perch diameter, clutter, and substrate texture and compliance. How many of these characteristics impact escape performance and/or escape behavior is poorly understood. However, the impact on locomotor performance has been identified for a few of these conditions.
Climbing up inclines not only poses a challenge for stability, but it also increases the cost of locomotion because a greater proportion of gravity acts to resist forward locomotion, leaving a smaller proportion of gravity to help hold the animal against the surface (Taylor et al. 1972; Cartmill 1985; Farley & Emshwiller 1996; Roberts et al. 1997; Preuschoft 2002; Daley & Biewener 2003; Autumn et al. 2006; Birn-Jeffery & Higham, 2014). This leads to a decrease in locomotor performance on steeper inclines. However, as discussed above, the magnitude of this decrease in performance is size dependent, permitting smaller animals to use inclines to facilitate escape from larger predators (Taylor et al. 1972; Irschick & Jayne 1999; Irschick 2003). One way to offset the challenges of moving up an incline is to increase muscle mass-specific power output. A recent examination of ten species of Anolis lizards found that the evolution toward a higher incidence of escaping upward (based on behavioral observation) has been paralleled by the evolution toward higher mass-specific power output (Vanhooydonck et al. 2006a). It is important to note that the degree of arboreality does not necessarily imply that a lizard will escape up an incline when confronted by a predator. In fact, some species of Anolis will escape up (e.g., A. valencienni), whereas others will not (e.g., A. lineatopus; Vanhooydonck et al. 2006a). Thus it is critical to match morphological and physiological measurements with natural behavior.
Narrower perches cause a reduction in locomotor performance primarily through their effects on stability. As substrate diameter decreases, so does the base of support, since foot placement is constrained to positions closer to the midline of the body. This increases the toppling moment about the perch, and thus decreases stability because a greater proportion of gravitational force acts tangentially to the perch surface (Cartmill 1985; Preuschoft 2002). The resulting decrease in locomotor performance (Losos & Sinervo 1989; Losos & Irschick 1996; Vanhooydonck et al. 2006b; but see Schmidt & Fischer 2010) presumably occurs because stability must be increased via postural changes (Figure 11.7; Peterson 1984; Higham & Jayne 2004b; Foster & Higham 2012), which, in turn, may affect muscle function (Foster & Higham 2014). As with increases in incline, smaller animals may benefit from retreating to a narrow perch during an escape, since larger animals would suffer a larger decrease in stability for a given perch diameter.
Despite these challenges, the arboreal habitat may offer some advantages in terms of refuge availability. The abundance of higher order branching and leaves, especially in the canopy, may obstruct locomotion by impeding limb movement, but it may also help reduce detectability of the prey by the predator, thereby reducing both frequency of escape events and necessary distance fled (Irschick & Garland 2001; Irschick 2003; Husak & Fox 2006). Further, obstruction due to clutter may also be size dependent, with larger predators unable to negotiate the barriers as effectively as smaller prey.
Complex branching patterns may also restrict either escape trajectory or escape behavior because a fleeing lizard will be forced to either execute sharp turns or jump to bridge gaps. In Anolis, locomotor speed decreases with increasing turning angle, regardless of escape strategy, because prey must pause to arrange limbs for jumping (Higham et al. 2001). However, the magnitude of this decrease differs among species (Higham et al. 2001).
The effect of perch compliance on lizard locomotion has been largely neglected. Its effect on jump performance in Anolis carolinensis suggests that increasing perch compliance decreases both take-off velocity and jump distance and alters jump trajectory due to recoil (Gilman et al. 2012). This species preferentially selects more rigid perches as take-off points for jumping in the field (Gilman & Irschick 2013). The effect of perch compliance on other locomotor behaviors and in other lizard species is unknown.
Finally, it is important to note that these challenges do not occur in isolation. Rather, fleeing lizards must encounter various combinations of these factors, some of which may have greater impacts on escape success than others. In some cases, the ability to rapidly alter locomotor behavior (through changes in kinematics) several times as various types of surfaces are encountered might determine the outcome of a predator-prey interaction. This ability to modulate locomotion, rather than the ability to move on any single perch, could be subject to strong selection. Although the relative importance of these challenges is poorly understood, Anolis lizards may select broader substrates, even if this requires sharper turning angles. This may reflect avoidance of instability caused by narrower surfaces to facilitate increased escape speeds (even if this reduces effective distance fled; Figure 11.7; Mattingly & Jayne 2005).
11.5.3 Saxicolous habitats
Many of the physical constraints of climbing described for arboreal lizards apply to saxicolous (rock-dwelling) lizards because steeply inclined surfaces are prevalent in rocky habitats. However, some rocky habitats also have the additional challenges of unsteady surfaces with variable rugosity (Figure 11.3; Revell et al. 2007 Goodman et al. 2008) that may lead to strong selection on locomotor morphology and performance (Taylor et al. 1972; Farley & Emshwiller 1996; Goodman et al. 2008; Collins et al. 2013). Interestingly, support for this hypothesis is equivocal. For example, whereas Liolaemus lizards exhibit no apparent ecomorphological or performance associations related to habitat use, Collins et al. (2013) found that saxicolous lizards, with longer tails, broader body shapes, and longer distal limb elements were less sensitive to substrate rugosity than their arboreal and psammophilous counterparts. Furthermore, in some lizards, saxicoly is tightly linked with increased jumping, clinging, and sprint performance (Goodman et al. 2008). The evolution of fast sprinting may be directly related to the distance an animal has to run to escape predation, which may be comparatively greater in rocky habitats than in other terrestrial habitats (Revell et al. 2007; Goodman 2009). Another possibility is that exposure to predators might be greater when a lizard is on top of a rock surface, as compared to being on an inclined branch of a tree (within clutter) or on the ground (with vegetation). However, conflicting hypotheses regarding how lizards deal with rocky habitats suggest that the three-dimensional structure of rocks, and how they are used during escape (e.g., jumping from rock to rock vs. climbing), may determine the morphological “fit” to the environment. Biomechanical studies, combined with corresponding behavioral studies in the field, would clarify how morphology is used under various circumstances.
Clinging and adhering are important in saxicolous habitats. In general, lizards with shorter, highly curved claws and short toes have a comparative clinging advantage relative to other lizards (Zani 2000). This is likely important for lizards that need to prevent detachment during predation attempts, though it does conflict with the need to attain high speeds or execute long jumps (Goodman et al. 2007, 2008; Collins et al. 2013). The toe pad of geckos is a key innovation that allows geckos to climb on the vertical and inverted surfaces of rocks and trees through friction and van der Waals interactions (Russell 1975, 1986; Autumn & Peattie 2002; Tian et al. 2006). Setae, the micro-structures comprising the toe pads, are hair-like structures ranging from 20 to 110 micrometers in length. Setae are pushed into the substrate and loaded in tension, thereby creating the intermolecular bonds that allow them to adhere (Autumn et al. 2000). If variation in micro-rugosity is high, only a percentage of setae may be able to engage with the surface at any given time, yet no evidence of morphological specialization for particular rugosities has been found. Instead, toe pads accommodate adhesion on a wide variety of unpredictable and highly variable structures (Russell & Johnson 2007, 2014). Collins et al. (2015) found that the morphology of the adhesive structures varies depending on the incline of the habitat. Those species that move on surfaces with relatively low inclines exhibit a reduction in the adhesive system.
11.5.4 Intermittent locomotion and habitat structure
As outlined above, habitat structure can profoundly impact escape behavior and performance. During an escape, pausing can be beneficial for avoiding fatigue and enhancing endurance in lizards (Weinstein & Full 1999), but can also be detrimental if it allows the predator to approach closer. Inclines and branching are two additional aspects of habitat structure that can increase intermittent locomotion (Higham et al. 2001, 2011b). On a vertical surface (compared to a level), Sceloporus woodi exhibits decreased running speeds, increased maximum acceleration, and increased pausing (Figure 11.9; Higham et al. 2011b). Thus it appears that either the enhanced acceleration may offset the costs associated with pausing or pausing facilitates increased acceleration by giving the propulsive muscles a chance to recover. With increasing turning angles, Anolis lizards pause more frequently to orient themselves before jumping across a turn (Higham et al. 2001), but these pauses may take valuable time during escapes, increasing the likelihood of predation. In addition to these potential benefits of intermittent locomotion, pausing may be beneficial for evaluating the continued need to escape, thus permitting an earlier return to activities such as foraging, courtship, and thermoregulation. Regardless, negotiating complex escape paths should be studied in greater detail because selection may act primarily on these demanding behaviors. We expect that there will be a trade-off between escaping into demanding areas of a habitat (steep inclines, narrow perches, areas with sharp turns, more compliant perches, etc.) and the ability to escape at high speeds, but we need to discover the context dependence of these strategies.
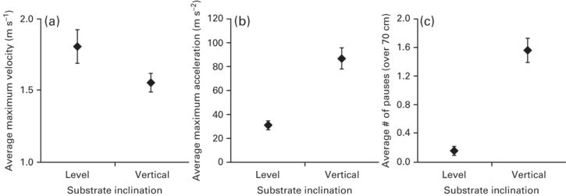
Figure 11.9
The average maximum velocity (A), acceleration (B), and number of pauses (C) in Sceloporus woodi moving along horizontal and vertical surfaces. Although the average acceleration was higher on the vertical surface compared to the horizontal surface, the average velocity was lower because the number of pauses was greater on the more energetically demanding vertical surface. However, greater acceleration in this condition may partially offset, and indeed may be possible because of, the effects of frequent pausing.
(Redrawn from Higham et al. 2011b)
11.6 Conclusions and future directions
The extraordinary diversity of behavior, morphology, and ecology among lizards makes them an ideal system for addressing many questions in biology, including those related to escape behavior and performance. Nevertheless, a number of facets contributing to lizard escape performance remain poorly understood.
Both temperature and motivation impact not only the decision of whether to escape, but the level of performance that is achieved during escape. To better understand how motivation affects the common measures of locomotor performance, we must improve our understanding of how such factors as personality traits, conflicting priorities (e.g., foraging, mating, defending territories), physiological state, and abiotic conditions contribute to motivation in ecologically relevant contexts (see also Careau & Garland 2012). Although a daunting prospect, such studies have the potential to profoundly affect our understanding of organismal biology.
Although performance measures may increase as motivation increases, the maximal level of performance during escape is limited by the prey’s locomotor ability. Numerous morphological and physiological traits interact to determine locomotor performance. Despite extensive research relating limb length and body size and shape to locomotor performance, there remains a paucity of data on the impact of differences in relative limb segment lengths and muscle morphology and physiology on locomotor capacity. Beyond this, however, it is essential that we establish the ecological relevance of such performance measures as acceleration and maximum sprint speed in order to ensure we are correctly characterizing and predicting the outcome of predator-prey interactions.
The impacts of hormones on locomotion in non-human vertebrates have received surprisingly little attention, despite their potential importance (O’Connor et al. 2011; Careau & Garland 2012; Higham & Irschick 2013). Although circulating levels of testosterone have been positively linked to increased performance in lizards (Husak et al. 2007), few significant impacts after experimentally elevating testosterone levels have been found. In the northern fence lizard, Sceloporus undulatus hyacinthinus, sprint speed and burst stamina were greater in individuals with experimentally elevated plasma testosterone (Klukowski et al. 1998). However, testosterone supplementation did not affect locomotor performance (burst distance and treadmill endurance) in Aspidoscelis sexlineata (O’Connor et al. 2011), leading to additional questions rather than answers. The conflicting results in the literature are likely a consequence of confounding factors associated with the experiments and natural differences among species and among populations within species (Crowley 1985a, b; Garland & Adolph 1991; Sinervo & Losos 1991; Bulova 1994; Snell et al. 1988; Van Damme et al. 1998; Iraeta et al. 2011).
Finally, lizards must interact with their habitat when escaping from predators. Thus the opportunities or challenges of aspects of substrates have the potential to impact performance capacity, resulting in the realized level of performance that the lizard achieves. However, the impact and relative importance of habitat characteristics on escape performance, rather than simply locomotor performance, is poorly understood. Biomechanical studies of ecologically relevant behaviors and interactions in the field are becoming increasingly feasible and are necessary to improve our understanding of escape performance in lizards.
Escaping from predators involves an integrated suite of behavioral, physiological, and morphological phenotypes, all of which may experience varying levels of selection in the wild. In addition, escape behavior and performance can be modulated depending on motivation. To fully understand when, how, and why lizards escape, future research should aim to integrate decision, motivation, ability, and opportunity. Only when we consider all of these categories simultaneously within a single lineage can we gain an appreciation for how they interact and interconnect to produce the observed escape performance and behavior.
References
Adams, B. (1985). The thermal-dependence of muscle membrane constants in 2 iguanid lizards (Dipsosaurus dorsalis, Sceloporus occidentalis). Federation Proceedings, 44, 1377-1377.
Aerts, P., Van Damme, R., Vanhooydonck, B., Zaaf, A. & Herrel, A. (2000). Lizard locomotion: How morphology meets ecology. Netherlands Journal of Zoology, 50, 261-277.
Alexander, R. M. (1977). Allometry of the limbs of antelopes (Bovidae). Journal of Zoology, 183, 125-146.
Allen, V., Elsey, R. M., Jones, N., Wright, J. & Hutchinson, J. R. (2010). Functional specialization and ontogenetic scaling of limb anatomy in Alligator mississippiensis. Journal of Anatomy, 216, 423-445.
Arnold, S. J. (1983). Morphology, performance and fitness. American Zoologist, 23, 347-361.
Astley, H. C., Abbott, E. M., Azizi, E., Marsh, R. L. & Roberts, T. J. (2013). Chasing maximal performance: a cautionary tale from the celebrated jumping frogs of Calaveras County. Journal of Experimental Biology, 216, 3947-3953.
Autumn, K. & Peattie, A. M. (2002). Mechanisms of adhesion in geckos. Integrative and Comparative Biology, 42, 1081-1090.
Autumn, K., Liang, Y. A., Hsieh, S. T., et al. (2000). Adhesive force of a single gecko foot-hair. Nature, 405, 681-685.
Autumn, K., Hsieh, S. T., Dudek, D. M., et al. (2006). Dynamics of geckos running vertically. Journal of Experimental Biology, 209, 260-272.
Bauer, A. M. & Russell, A. P. (1991). Pedal specializations in dune-dwelling geckos. Journal of Arid Environments, 20, 43-62.
Beck, R. C. (1978). Motivation: Theories and Principles. Englewood Cliffs: Prentice-Hall, Inc.
Bellairs, A. d. A. (1970). The Life of Reptiles. New York: Universe Books.
Bellairs, A. d. A. & Bryant, S. V. (1985). Autotomy and regeneration in reptiles. In Gans, C. & Billett, F. (eds.) Biology of the Reptilia. New York: John Wiley and Sons, 15, 301-410.
Bennett, A. F. & Huey, R. B. (1990). Studying the evolution of physiological performance. In Futuyma, D. J. & Antonovics, J. (eds.) Oxford Surveys in Evolutionary Biology, Vol. 6. Oxford: Oxford University Press, pp. 251-284.
Bergmann, P. J., Meyers, J. J. & Irschick, D. J. (2009). Directional evolution of stockiness coevolves with ecology and locomotion in lizards. Evolution, 63, 215.
Biewener, A. A. (1998). Muscle function in vivo: A comparison of muscles used for elastic energy savings versus muscles used to generate mechanical power. American Zoologist, 38, 703-717.
Biewener, A. A. & Roberts, T. J. (2000). Muscle and tendon contributions to force, work, and elastic energy savings: a comparative perspective. Exercise and Sport Sciences Reviews, 28, 99-107.
Birn-Jeffery, A. & Higham, T. E. (2014). The scaling of uphill and downhill locomotion in legged animals. Integrative and Comparative Biology, 54, 1159-1172.
Bonine, K. E. & Garland, T., Jr. (1999). Sprint performance of phrynosomatid lizards, measured on a high-speed treadmill, correlates with hindlimb length. Journal of Zoology, 248, 255-265.
Bonine, K. E., Gleeson, T. T. & Garland, T., Jr. (2001). Comparative analysis of fiber-type composition in the iliofibularis muscle of phrynosomatid lizards (Squamata). Journal of Morphology, 250, 265-280.
Bonine, K. E., Gleeson, T. T. & Garland, T., Jr. (2005). Muscle fiber-type variation in lizards (Squamata) and phylogenetic reconstruction of hypothesized ancestral states. Journal of Experimental Biology, 208, 4529-4547.
Brainerd, E. L. & Owerkowicz, T. (2006). Functional morphology and evolution of aspiration breathing in tetrapods. Respiratory Physiology & Neurobiology, 154, 73-88.
Brainerd, E. L. & Patek, S. N. (1998). Vertebral column morphology, C-start curvature, and the evolution of mechanical defenses in tetraodontiform fishes. Copeia, 1998, 971-984.
Brodie, E. D., III. (1989). Behavioral modification as a means of reducing the cost of reproduction. American Naturalist, 134, 225-238.
Bulova, S. J. (1994). Ecological correlates of population and individual variation in antipredator behavior of two species of desert lizards. Copeia, 1994, 980-992.
Careau, V. & Garland, T., Jr. (2012). Performance, personality, and energetics: Correlation, causation, and mechanism. Physiological and Biochemical Zoology, 85, 543-571.
Careau, V., Wolak, M. E., Carter, P. A. & Garland, T., Jr. (2013). Limits to behavioral evolution: The quantitative genetics of a complex trait under directional selection. Evolution, 67, 3102-3119.
Carothers, J. H. (1986). An experimental confirmation of morphological adaptation: Toe fringes in the sand-dwelling lizard Uma scoparia. Evolution, 40, 871-874.
Carrier, D. R. (1991). Conflict in the hypaxial musculo-skeletal system: Documenting an evolutionary constraint. American Zoologist, 31, 644-654.
Carrier, D. R. (1996). Ontogenetic limits on locomotor performance. Physiological Zoology, 69, 467-488.
Cartmill, M. (1985). Climbing. In Hildebrand, M., Bramble, D. M., Liem, K. F. & Wake, D. B. (eds.) Functional Vertebrate Morphology. Cambridge: Harvard University Press, pp. 73-88.
Christian, A. & Garland, T., Jr. (1996). Scaling of limb proportions in monitor lizards (Squamata: Varanidae). Journal of Herpetology, 30, 219-230.
Clark, D. R., Jr. (1971). The strategy of tail-autotomy in the ground skink, Lygosoma laterale. Journal of Experimental Zoology, 176, 295-302.
Clemente, C. J., Withers, P. C., Thompson, G., & Loyd, D. (2013). Lizard tricks: Overcoming conflicting requirements of speed versus climbing ability by altering biomechanics of the lizard stride. Journal of Experimental Biology, 216, 3854-3862.
Collar, D. C., Schulte, J. A., II, & Losos, J. B. (2011). Evolution of extreme body size disparity in monitor lizards (Varanus). Evolution, 65, 2664-2680.
Collins, C. E., Russell, A. P. & Higham, T. E. (2015). Subdigital adhesive pad morphology varies in relation to structural habitat use in the Namib Day Gecko, Rhoptropus afer. Functional Ecology. 29, 66-77.
Collins, C. E., Self, J. D., Anderson, R. A. & McBrayer, L. D. (2013). Rock-dwelling lizards exhibit less sensitivity of sprint speed to increases in substrate rugosity. Zoology, 116, 151-158.
Coombs, W. P., Jr. (1978). Theoretical aspects of cursorial adaptations in dinosaurs. Quarterly Review of Biology, 53, 393-418.
Cooper, W. E., Jr. (2000). Effect of temperature on escape behaviour by an ectothermic vertebrate, the keeled earless lizard (Holbrookia propinqua). Behaviour, 137, 1299-1315.
Cooper, W. E., Jr. (2008). Visual monitoring of predators: Occurrence, cost and benefit for escape. Animal Behaviour, 76, 1365-1372.
Cooper, W. E., Jr. & Frederick, W. G. (2007). Optimal flight initiation distance. Journal of Theoretical Biology, 244, 59-67.
Cooper, W. E., Jr. & Vitt, L. J. (1991). Influence of detectability and ability to escape on natural selection of conspicuous autonomous defenses. Canadian Journal of Zoology, 69, 757-764.
Cooper, W. E., Jr. & Wilson, D. S. (2007). Beyond optimal escape theory: Microhabitats as well as predation risk affect escape and refuge use by the phrynosomatid lizard Sceloporus virgatus. Behaviour, 144, 1235-1254.
Cooper, W. E., Jr., Vitt, L. J., Hedges, R. & Huey, R. B. (1990). Locomotor impairment and defense in gravid lizards (Eumeces laticeps): Behavioral shift in activity may offset costs of reproduction in an active forager. Behavioral Ecology and Sociobiology, 27, 153-157.
Cooper, W. E., Jr., Pyron, R. A. & Garland, T., Jr. (2014). Island tameness: Living on islands reduces flight initiation distance. Proceedings of the Royal Society B, 281, 20133019.
Crowley, S. R. (1985a). Thermal sensitivity of sprint-running in the lizard Sceloporus undulatus: support for a conservative view of thermal physiology. Oecologia, 66, 219-225.
Crowley, S. R. (1985b). Insensitivity to desiccation of sprint running performance in the lizard, Sceloporus undulatus. Journal of Herpetology, 19, 171-174.
Crowley, S. R. & Pietruszka, R. D. (1983). Aggressiveness and vocalization in the leopard lizard. (Gambelia wislizennii): The influence of temperature. Animal Behaviour, 31, 1055-1060.
Daley, M. A. & Biewener, A. A. (2003). Muscle force-length dynamics during level versus incline locomotion: A comparison of in vivo performance of two guinea fowl ankle extensors. Journal of Experimental Biology, 206, 2941-2958.
Daniels, C. B. (1983). Running: An escape strategy enhanced by autotomy. Herpetologica, 39, 162-165.
Daniels, C. B., Flaherty, S. P. & Simbotwe, M. P. (1986). Tail size and effectiveness of autotomy in a lizard. Journal of Herpetology, 20, 93-96.
Dickinson, M. H., Farley, C. T., Full, R. J., et al. (2000). How animals move: An integrative view. Science, 288, 100-106.
Djawdan, M. (1993). Locomotor performance of bipedal and quadrupedal heteromyid rodents. Functional Ecology, 7, 195-202.
Farley, C. & Emshwiller, M. (1996). Efficiency of uphill locomotion in nocturnal and diurnal lizards. Journal of Experimental Biology, 199, 587-592.
Farmer, C. G. & Carrier, D. R. (2000a). Pelvic aspiration in the American alligator (Alligator mississippiensis). Journal of Experimental Biology, 203, 1679-1687.
Farmer, C. G. & Carrier, D. R. (2000b). Ventilation and gas exchange during treadmill locomotion in the American alligator (Alligator mississippiensis). Journal of Experimental Biology, 203, 1671-1678.
Feder, M. E., Garland, T., Jr., Marden, J. H. & Zera, A. J. (2010). Locomotion in response to shifting climate zones: Not so fast. Annual Review of Physiology, 72, 167-190.
Fitts, R. H., McDonald, K. S. & Schluter, J. M. (1991). The determinants of skeletal muscle force and power: Their adaptability with changes in activity pattern. Journal of Biomechanics, 24, 111-122.
Foster, K. L. & Higham, T. E. (2012). How fore- and hindlimb function changes with incline and perch diameter in the green anole, Anolis carolinensis. Journal of Experimental Biology, 215, 2288-2300.
Foster, K. L. & Higham, T. E. (2014). Context-dependent changes in motor control and kinematics during locomotion: Modulation and decoupling. Proceedings of the Royal Society B, 281, 20133331.
Gans, C. & de Vree, F. (1987). Functional bases of fiber length and angulation in muscle. Journal of Morphology, 192, 63-85.
Garland, T., Jr. (1985). Ontogenetic and individual variation in size, shape and speed in the Australian agamid lizard Amphibolurus nuchalis. Journal of Zoology, 207, 425-439.
Garland, T., Jr. (1993). Locomotor performance and activity metabolism of Cnemidophorus tigris in relation to natural behaviors. In Wright, J. W. & Vitt, L. J. (eds.) Biology of Whiptail Lizards (Genus Cnemidophorus). Norman: Oklahoma Museum of Natural History, pp. 163-210.
Garland, T., Jr. (1994). Phylogenetic analyses of lizard endurance capacity in relation to body size and body temperature. In Vitt, L. J. & Pianka, E. R. (eds.) Lizard Ecology: Historical and Experimental Perspectives. Princeton: Princeton University Press, pp. 237-259.
Garland, T., Jr. (2014). Quick guide: Trade-offs. Current Biology, 24, R60-R61.
Garland, T., Jr. & Adolph, S. C. (1991). Physiological differentiation of vertebrate populations. Annual Review of Ecology and Systematics, 22, 193-228.
Garland, T., Jr. & Else, P. L. (1987). Seasonal, sexual, and individual variation in endurance and activity metabolism in lizards. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 252, R439-R449.
Garland, T., Jr. & Janis, C. M. (1993). Does metatarsal/femur ratio predict maximal running speed in cursorial mammals? Journal of Zoology, 229, 133-151.
Garland, T., Jr. & Losos, J. B. (1994). Ecological morphology of locomotor performance in squamate reptiles. In Wainwright, P. C. & Reilly, S. M. (eds.) Ecological Morphology: Integrative Organismal Biology. Chicago: University of Chicago Press, pp. 240-302.
Garland, T., Jr., Kelly, S. A., Malisch, J. L. et al. (2011). How to run far: Multiple solutions and sex-specific responses to selective breeding for high voluntary activity levels. Proceedings of the Royal Society B: Biological Sciences, 278, 574-581.
Gilbert, E. A. B., Payne, S. L. & Vickaryous, M. K. (2013). The anatomy and histology of caudal autotomy and regeneration in lizards. Physiological & Biochemical Zoology, 86, 631-644.
Gillis, G. B., Bonvini, L. A. & Irschick, D. J. (2009). Losing stability: Tail loss and jumping in the arboreal lizard Anolis carolinensis. Journal of Experimental Biology, 212, 604-609.
Gillis, G. B., Kuo, C.-Y. & Irschick, D. J. (2013). The impact of tail loss on stability during jumping in green anoles (Anolis carolinensis). Physiological & Biochemical Zoology, 86, 680-689.
Gilman, C. A. & Irschick, D. J. (2013). Foils of flexion: The effects of perch compliance on lizard locomotion and perch choice in the wild. Functional Ecology, 27, 374-381.
Gilman, C. A., Bartlett, M. D., Gillis, G. B. & Irschick, D. J. (2012). Total recoil: Perch compliance alters jumping performance and kinematics in green anole lizards (Anolis carolinensis). Journal of Experimental Biology, 215, 220-226.
Glasheen, J. W. & McMahon, T. A. (1996). Size-dependence of water-running ability in basilisk lizards (Basiliscus basiliscus). Journal of Experimental Biology, 199, 2611-2618.
Gleeson, T. T. & Harrison, J. M. (1988). Muscle composition and its relation to sprint running in the lizard Dipsosaurus dorsalis. American Journal of Physiology, 255, R470-R477.
Goodman, B. A. (2007). Divergent morphologies, performance, and escape behaviour in two tropical rock-using lizards (Reptilia: Scincidae). Biological Journal of the Linnean Society, 91, 85-98.
Goodman, B. A. (2009). Nowhere to run: The role of habitat openness and refuge use in defining patterns of morphological and performance evolution in tropical lizards. Journal of Evolutionary Biology, 22, 1535-1544.
Goodman, B. A., Miles, D. B. & Schwarzkopf, L. (2008). Life on the rocks: Habitat use drives morphological and performance evolution in lizards. Ecology, 89, 3462-3471.
Hansen, W. R. & Autumn, K. (2005). Evidence for self-cleaning in Gecko setae. Proceedings of the National Academy of Sciences, 102, 385-389.
Haxton, H. A. (1944). Absolute muscle force in the ankle flexors of man. Journal of Physiology, 103, 267-273.
Herrel, A., James, R. S. & Van Damme, R. (2007). Fight versus flight: Physiological basis for temperature-dependent behavioral shifts in lizards. Journal of Experimental Biology, 210, 1762-1767.
Herrel, A., Vanhooydonck, B., Porck, J. & Irschick, D. J. (2008). Anatomical basis of differences in locomotor behavior in Anolis lizards: A comparison between two ecomorphs. Bulletin of the Museum of Comparative Zoology, 159, 213-238.
Hertz, P. E., Huey, R. B. & Nevo, E. (1982). Fight versus flight: Body temperature influences defensive responses of lizards. Animal Behaviour, 30, 676-679.
Hertz, P. E., Huey, R. B. & Garland, T., Jr. (1988). Time budgets, thermoregulation, and maximal locomotor performance: Are reptiles olympians or boy scouts? American Zoologist, 28, 927-938.
Herzog, W. (2000). Muscle properties and coordination during voluntary movement. Journal of Sports Science, 18, 141-152.
Higham, T. E. & Anderson, C. V. (2013). Function and adaptation. In Tolley, K. A. & Herrel, A. (eds.) The Biology of Chameleons. Berkeley: University of California Press, pp. 63-83.
Higham, T. E. & Irschick, D. J. (2013). Springs, steroids, and slingshots: The roles of enhancers and constraints in animal movement. Journal of Comparative Physiology B, 183, 583-595.
Higham, T. E. & Jayne, B. C. (2004a). In vivo muscle activity in the hindlimb of the arboreal lizard, Chamaeleo calyptratus: General patterns and the effects of incline. Journal of Experimental Biology, 207, 249-261.
Higham, T. E. & Jayne, B. C. (2004b). Locomotion of lizards on inclines and perches: Hindlimb kinematics of an arboreal specialist and a terrestrial generalist. Journal of Experimental Biology, 207, 233-248.
Higham, T. E. & Russell, A. P. (2010). Flip, flop and fly: Modulated motor control and highly variable movement patterns of autotomized gecko tails. Biology Letters, 6, 70-73.
Higham, T. E. & Russell, A. P. (2012). Time-varying motor control of autotomized leopard gecko tails: Multiple inputs and behavioral modulation. Journal of Experimental Biology, 215, 435-441.
Higham, T. E., Davenport, M. S. & Jayne, B. C. (2001). Maneuvering in an arboreal habitat: The effects of turning angle on the locomotion of three sympatric ecomorphs of Anolis lizards. Journal of Experimental Biology, 204, 4141-4155.
Higham, T. E., Korchari, P. G. & McBrayer, L. M. (2011a). How muscles define maximum locomotor performance in lizards: An analysis using stance and swing phase muscles. Journal of Experimental Biology, 214, 1685-1691.
Higham, T. E., Korchari, P. G. & McBrayer, L. M. (2011b). How to climb a tree: Lizards accelerate faster, but pause more, when escaping on vertical surfaces. Biological Journal of the Linnean Society, 102, 83-90.
Higham, T. E., Lipsett, K. R., Syme, D. A. & Russell, A. P. (2013a). Controlled chaos: Three-dimensional kinematics, fiber histochemistry and muscle contractile dynamics of autotomized lizard tails. Physiological and Biochemical Zoology, 86, 611-630.
Higham, T. E., Russell, A. P. & Zani, P. A. (2013b). Integrative biology of tail autotomy in lizards. Physiological and Biochemical Zoology, 86, 603-610.
Hildebrand, M. (1985). Walking and running. In Hildebrand, M., Bramble, D. M., Liem, K.F. & Wake, D. B. (eds.) Functional Vertebrate Morphology. Cambridge: Harvard University Press, pp. 38-57.
Howland, H. C. (1974). Optimal strategies for predator avoidance: The relative importance of speed and manoeuvrability. Journal of Theoretical Biology, 47, 333-350.
Hsieh, S. T. & Lauder, G. V. (2004). Running on water: Three-dimensional force generation by basilisk lizards. Proceedings of the National Academy of Sciences, 101, 16784-16788.
Huey, R. B. & Dunham, A. E. (1987). Repeatability of locomotor performance in natural populations of the lizard Sceloporus merriami. Evolution, 41, 1116-1120.
Huey, R. B. & Hertz, P. E. (1984). Effects of body size and slope on acceleration of a lizard (Stellio stellio). Journal of Experimental Biology, 110, 113-123.
Huey, R. B., Dunham, A. E., Overall, K. L. & Newman, R. A. (1990). Variation in locomotor performance in demographically known populations of the lizard Sceloporus merriami. Physiological Zoology, 63, 845-872.
Husak, J. F. (2006a). Does speed help you survive? A test with collared lizards of different ages. Functional Ecology, 20, 174-179.
Husak, J. F. (2006b). Does survival depend on how fast you can run or how fast you do run? Functional Ecology, 20, 1080-1086.
Husak, J. F. (2006c). Do female collared lizards change field use of maximal sprint speed capacity when gravid? Oecologia, 150, 339-343.
Husak, J. F. & Fox, S. F. (2006). Field use of maximal sprint speed by collared lizards (Crotaphytus collaris): Compensation and sexual selection. Evolution, 60, 1888-1895.
Husak, J. F. & Fox, S. F. (2008). Sexual selection on locomotor performance. Evolutionary Ecology Research, 10, 213-228.
Husak, J. F., Irschick, D. J., Meyers, J. J., Lailvaux, S. P. & Moore, I. T. (2007). Hormones, sexual signals, and performance of green anole lizards (Anolis carolinensis). Hormones and Behavior, 52, 360-367.
Iraeta, P., Monasterio, C., Salvador, A. & Díaz, J. A. (2011). Sexual dimorphism and interpopulation differences in lizard hind limb length: Locomotor performance or chemical signalling? Biological Journal of the Linnean Society, 104, 318-329.
Irschick, D. J. (2000). Effects of behaviour and ontogeny on the locomotor performance of a West Indian lizard, Anolis lineatopus. Functional Ecology, 14, 438-444.
Irschick, D. J. (2003). Measuring performance in nature: implications for studies of fitness within populations. Integrative and Comparative Biology, 43, 396-407.
Irschick, D. J. & Garland, T. (2001). Integrating function and ecology in studies of adaptation: Investigations of locomotor capacity as a model system. Annual Review of Ecology and Systematics, 32, 367-396.
Irschick, D. J. & Jayne, B. C. (1999). A field study of the effects of incline on the escape locomotion of a bipedal lizard, Callisaurus draconoides. Physiological & Biochemical Zoology, 72, 44-56.
Irschick, D. J. & Losos, J. B. (1998). A comparative analysis of the ecological significance of maximal locomotor performance in Caribbean Anolis lizards. Evolution, 52, 219-226.
Irschick, D. J. & Losos, J. B. (1999). Do lizards avoid habitats in which performance is submaximal? The relationship between sprinting capabilities and structural habitat use in Caribbean anoles. American Naturalist, 154, 293-305.
Irschick, D. J., Vanhooydonck, B., Herrel, A. & Andronescu, A. (2003). Effects of loading and size on maximum power output and gait characteristics in geckos. Journal of Experimental Biology, 206, 3923-3934.
Irschick, D. J., Herrel, A., Vanhooydonck, B., Huyghe, K. & van Damme, R. (2005). Locomotor compensation creates a mismatch between laboratory and field estimates of escape speed in lizards: A cautionary tale for performance-to-fitness studies. Evolution, 59, 1579-1587.
Jaksić, F. M. & Núñez, H. (1979). Escaping behavior and morphological correlates in two Liolaemus species of central Chile (Lacertilia: Iguanidae). Oecologia, 42, 119-122.
James, R. S. (2013). A review of the thermal sensitivity of the mechanics of vertebrate skeletal muscle. Journal of Comparative Physiology B, 183, 723-733.
Jayne, B. C. & Daggy, M. W. (2000). The effects of temperature on the burial performance and axial motor pattern of the sand-swimming of the Mojave fringe-toed lizard Uma scoparia. Journal of Experimental Biology, 203, 1241-1252.
Jayne, B. C. & Ellis, R. V. (1998). How inclines affect the escape behaviour of a dune-dwelling lizard, Uma scoparia. Animal Behavior, 55, 1115-1130.
Jayne, B. C. & Irschick, D. J. (1999). Effects of incline and speed on the three-dimensional hindlimb kinematics of a generalized iguanian lizard (Dipsosaurus dorsalis). Journal of Experimental Biology, 202, 143-159.
Jayne, B. C. & Irschick, D. J. (2000). A field study of incline use and preferred speeds for the locomotion of lizards. Ecology, 81, 2969-2983.
John-Alder, H. B., Garland, T., Jr. & A. F., Bennett, (1986). Locomotory capacities, oxygen consumption, and the cost of locomotion of the shingle-back lizard (Trachydosaurus rugosus). Physiological Zoology, 59, 523-531.
Klukowski, M., Jenkinson, N. M. & Nelson, C. E. (1998). Effects of testosterone on locomotor performance and growth in field-active northern fence lizards, Sceloporus undulatus hyacinthinus. Physiological Zoology, 71, 506-514.
Kohlsdorf, T. & Navas, C. (2012). Evolution of form and function: Morphophysiological relationships and locomotor performance in tropidurine lizards. Journal of Zoology, 288, 41-49.
Lailvaux, S. P. (2007). Interactive effects of sex and temperature on locomotion in reptiles. Integrative and Comparative Biology, 47, 189-199.
Lailvaux, S. P., Alexander, G. J. & Whiting, M. J. (2003). Sex-based differences and similarities in locomotor performance, thermal preferences, and escape behaviour in the lizard Platysaurus intermedius wilhelmi. Physiological & Biochemical Zoology, 76, 511-521.
Lamb, T. & Aaron, M. B. (2006). Footprints in the sand: Independent reduction of subdigital lamellae in the Namib-Kalahari burrowing geckos. Proceedings of the Royal Society, B., 273, 855-864.
Le Galliard, J. F., Le Bris, M. & Clobert, J. (2003). Timing of locomotor impairment and shift in thermal preferences during gravidity in a viviparous lizard. Functional Ecology, 17, 877-885.
Lejeune, T. M., Willems, P. A. & Heglund, N. C. (1998). Mechanics and energetics of human locomotion on sand. Journal of Experimental Biology, 201, 2071-2080.
Li, C., Hsieh, S. T. & Goldman, D. I. (2012). Multi-functional foot use during running in the zebra-tailed lizard (Callisaurus draconoides). Journal of Experimental Biology, 215, 3293-3308.
Lieber, R. L. & Ward, S. R. (2011). Skeletal muscle design to meet functional demands. Philosophical Transactions of the Royal Society B, 366, 1466-1476.
Loeb, G. E. & Gans, C. (1986). The organization of muscle. In Electromyography for Experimentalists. London: University of Chicago Press, pp. 25-43.
Losos, J. B. & Irschick, D. J. (1996). The effect of perch diameter on escape behaviour of Anolis lizards: Laboratory predictions and field tests. Animal Behaviour, 51, 593-602.
Losos, J. B. & Sinervo, B. (1989). The effects of morphology and perch diameter on sprint performance of Anolis lizards. Journal of Experimental Biology, 145, 23-30.
Luke, C. (1986). Convergent evolution of lizard toe fringes. Biological Journal of the Linnean Society, 27, 1-16.
Maladen, R. D., Ding, Y., Li, C. & Goldman, D. I.(2009). Undulatory swimming in sand: Subsurface locomotion of the sandfish lizard. Science, 325, 314-318.
Marsh, R. L. & Bennett, A. F. (1986a). Thermal-dependence of contractile properties of skeletal-muscle from the lizard Sceloporus occidentalis with comments on methods for fitting and comparing force-velocity curves. Journal of Experimental Biology, 126, 63-77.
Marsh, R. L. & Bennett, A. F. (1986b). Thermal-dependence of sprint performance of the lizard Sceloporus occidentalis. Journal of Experimental Biology, 126, 79-87.
Mattingly, W. B. & Jayne, B. C. (2005). The choice of arboreal escape paths and its consequences for the locomotor behaviour of four species of Anolis lizards. Animal Behaviour, 70, 1239-1250.
McElroy, E. & Bergmann, P. J. (2013). Tail autotomy, tail size and locomotor performance in lizards. Physiological & Biochemical Zoology, 86, 669-679.
McElroy, E. J. & McBrayer, L. D. (2010). Getting up to speed: Acceleration strategies in the Florida scrub lizard, Sceloporus woodi. Physiological & Biochemical Zoology, 83, 643-653.
McElroy, E. J., Meyers, J. J., Reilly, S. M. & Irschick, D. J. (2007). Dissecting the effects of behaviour and habitat on the locomotion of a lizard (Urosaurus ornatus). Animal Behaviour, 73, 359-365.
McGuire, J. A. (2003). Allometric prediction of locomotor performance: An example from Southeast Asian flying lizards. American Naturalist, 161, 337-349.
Melville, J. (2008). Evolutionary correlations between microhabitat specialisation and locomotor capabilities in the lizard genus Niveoscincus. Australian Journal of Zoology, 55, 351-355.
Melville, J. & Swain, R. (2000). Evolutionary relationships between morphology, performance and habitat openness in the lizard genus Niveoscincus (Scincidae : Lygosominae). Biological Journal of the Linnean Society, 70, 667-683.
O’Connor, J. L., McBrayer, L. M., Higham, T. E. et al. (2011). Effects of training and testosterone on muscle fiber types and locomotor performance in male six-lined racerunners (Aspidoscelis sexlineata). Physiological and Biochemical Zoology, 84, 394-405.
Payne, R. C., Crompton, R. H., Isler, K., et al. (2006). Morphological analysis of the hindlimb in apes and humans. II. Moment arms. Journal of Anatomy, 208, 725-742.
Peterson, J. A. (1984). The locomotion of Chamaeleo (Reptilia: Sauria) with particular reference to the forelimb. Journal of Zoology, 202, 1-42.
Pianka, E. R. & Vitt, L. J. (2003). Lizards: Windows to the Evolution of Diversity. Berkeley, CA: University of California Press.
Preuschoft, H. (2002). What does “arboreal locomotion” mean exactly and what are the relationships between “climbing”, environment and morphology? Zeitschrift fur Morphologie und Anthropologie, 83, 171-188.
Rand, A. S. (1964). Ecological distribution in anoline lizards of Puerto Rico. Ecology, 45, 745-752.
Rassier, D. E., MacIntosh, B. R. & Herzog, W. (1999). Length dependence of active force production in skeletal muscle. Journal of Applied Physiology, 86, 1445-1457.
Reilly, S. M. (1998). Sprawling locomotion in the lizard Sceloporus clarkii: speed modulation of motor patterns in a walking trot. Brain, Behavior & Evolution, 52, 126-138.
Reilly, S. M. & Delancey, M. J. (1997). Sprawling locomotion in the lizard Sceloporus clarkii: the effects of speed on gait, hindlimb kinematics, and axial bending during walking. Journal of Zoology, 243, 417-433.
Revell, L. J., Johnson, M. A., Schulte, J. A., II, Kolbe, J. J. & Losos, J. B. (2007). A phylogenetic test for adaptive convergence in rock-dwelling lizards. Evolution, 61, 2898-2912.
Reznick, D. (1985). Costs of reproduction: An evaluation of the empirical evidence. Oikos, 44, 257-267.
Richmond, F. J. R. (1998). Elements of style in neuromuscular architecture. American Zoologist, 38, 729-742.
Roberts, T. J., Marsh, R. L., Weyand, P. G. & Taylor, C. R. (1997). Muscular force in running turkeys: the economy of minimizing work. Science, 275, 1113-1115.
Rome, L. C. & Bennett, A. F. (1990). Influence of temperature on muscle and locomotor performance. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 259, R189-R190.
Runyon, D. (1992). Guys and Dolls. New York: Penguin Books, Ltd.
Russell, A. P. (1975). A contribution to the functional analysis of the foot of the Tokay, Gekko gecko (Reptilia: Gekkonidae). Journal of Zoology, 176, 437-476.
Russell, A. P. (1986). The morphological basis of weight-bearing in the scansors of the tokay gecko (Reptilia: Sauria). Canadian Journal of Zoology, 64, 948-955.
Russell, A. P. & Johnson, M. K. (2007). Real-world challenges to, and capabilities of, the gekkotan adhesive system: contrasting the rough and the smooth. Canadian Journal of Zoology, 85, 1228-1238.
Russell, A. P. & Johnson, M. K. (2014). Between a rock and a soft place: Microtopography of the locomotor substrate and the morphology of the setal fields of Namibian day geckos (Gekkota: Gekkonidae: Rhoptropus). Acta Zoologica, 95, 295-318.
Russell, A. P., Lai, E. K., Powell, G. L. & Higham, T. E. (2014). Density and distribution of cutaneous sensilla on tails of leopard geckos (Eublepharis macularius) in relation to caudal autotomy. Journal of Morphology, 275, 961-979.
Sacks, R. D. & Roy, R. R. (1982). Architecture of the hind limb muscles of cats: Functional significance. Journal of Morphology, 173, 185-195.
Scales, J. & Butler, M. (2007). Are powerful females powerful enough? Acceleration in gravid green iguanas (Iguana iguana). Integrative and Comparative Biology, 47, 285-294.
Schall, J. J. & Pianka, E. R. (1980). Evolution of escape behavior diversity. American Naturalist, 115, 551-566.
Schmidt, A. & Fischer, M. S. (2010). Arboreal locomotion in rats. The challenge of maintaining stability. Journal of Experimental Biology, 213, 3615-3624.
Sherbrooke, W. C., George, A. & Middendorf, G. A., III. (2001). Blood-squirting variability in horned lizards (Phrynosoma). Copeia, 2001, 1114-1122.
Shine, R. (2003a). Effects of pregnancy on locomotor performance: An experimental study on lizards. Oecologia, 136, 450-456.
Shine, R. (2003b). Locomotor speeds of gravid lizards: Placing “costs of reproduction” within an ecological context. Functional Ecology, 17, 526-533.
Sinervo, B. & Losos, J. B.(1991). Walking the tight rope: arboreal sprint performance among Sceloporus occidentalis lizard populations. Ecology, 72, 1225-1233.
Skelton, T. M., Waran, N. K. & Young, R. J. (1996). Assessment of motivation in the lizard, Chalcides ocellatus. Animal Welfare, 5, 63-69.
Smith, D. G. (1997). Ecological factors influencing the antipredator behaviors of the ground skink, Scincella lateralis. Behavioral Ecology, 8, 622-629.
Snell, H. L., Jennings, R. D., Snell, H. M. & Harcourt, S. (1988). Intrapopulation variation in predator-avoidance performance of Galápagos lava lizards: The interaction of sexual and natural selection. Evolutionary Ecology, 2, 353-369.
Snyder, R. C. (1954). The anatomy and function of the pelvic girdle and hindlimb in lizard locomotion. American Journal of Anatomy, 95, 1-45.
Snyder, R. C. (1962). Adaptations for bipedal locomotion of lizards. American Zoologist, 2, 191-203.
Sorci, G., Swallow, J. G., Theodore, G., Jr. & Clobert, J. (1995). Quantitative genetics of locomotor speed and endurance in the lizard Lacerta vivipara. Physiological Zoology, 68, 698-720.
Spezzano, L. C. & Jayne, B. C. (2004). The effects of surface diameter and incline on the hindlimb kinematics of an arboreal lizard (Anolis sagrei). Journal of Experimental Biology, 207, 2115-2131.
Stiller, R. B. & McBrayer, L. D. (2013). The ontogeny of escape behavior, locomotor performance, and the hind limb in Sceloporus woodi. Zoology, 116, 175-181.
Taylor, C. R., Caldwell, S. L. & Rowntree, V. J. (1972). Running up and down hills: Some consequences of size. Science, 178, 1096-1097.
Tian, Y., Pesika, N., Zeng, H. et al. (2006). Adhesion and friction in gecko toe attachment and detachment. Proceedings of the National Academy of Sciences, 103, 19320-19325.
Toro, E., Herrel, A., Vanhooydonck, B. & Irschick, D. J. (2003). A biomechanical analysis of intra- and interspecific scaling of jumping and morphology in Caribbean Anolis lizards. Journal of Experimental Biology, 206, 2641-2652.
Toro, E., Herrel, A. & Irschick, D. J. (2004). The evolution of jumping performance in Caribbean Anolis lizards: Solutions to biomechanical trade-offs. American Naturalist, 163, 844-856.
Tulli, M. J., Cruz, F. B., Herrel, A., Vanhooydonck, B. & Abdala, V. (2009). The interplay between claw morphology and microhabitat use in neotropical iguanian lizards. Zoology, 112, 379-392.
Tulli, M. J., Abdala, V. & Cruz, F. B. (2012). Effects of different substrates on the sprint performance of lizards. Journal of Experimental Biology, 215, 774-784.
Van Damme, R., Aerts, P. & Vanhooydonck, B. (1998). Variation in morphology, gait characteristics and speed of locomotion in two populations of lizards. Biological Journal of the Linnean Society, 63, 409-427.
Vanhooydonck, B., Aerts, P., Irschick, D.J. & Herrel, A. (2006a). Power generation during locomotion in Anolis lizards: An ecomorphological approach. In Herrel, A., Speck, T. & Rowe, N.P. (eds.) Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants. Boca Raton, FL: CRC Press, pp. 253-269.
Vanhooydonck, B., Herrel, A. & Irschick, D. J. (2006b). Out on a limb: the differential effect of substrate diameter on acceleration capacity in Anolis lizards. Journal of Experimental Biology, 209, 4515-4523.
Vanhooydonck, B., Herrel, A., Van Damme, R. & Irschick, D. J. (2006c). The quick and the fast: The evolution of acceleration capacity in Anolis lizards. Evolution, 60, 2137-2147.
Vanhooydonck, B., James, R. S., Tallis, J., et al. (2014). Is the whole more than the sum of its parts? Evolutionary trade-offs between burst and sustained locomotion in lacertid lizards. Proceedings of the Royal Society B: Biological Sciences, 281, 20132677.
Wainwright, P. C. (2007). Functional versus morphological diversity in macroevolution. Annual Review of Ecology, Evolution, and Systematics, 38, 381-401.
Wainwright, P. C., Alfaro, M. E., Bolnick, D. I. & Hulsey, C. D. (2005). Many-to-one mapping of form to function: A general principle in organismal design? Integrative and Comparative Biology, 45, 256-262.
Walker, J. A. (2000). Does a rigid body limit maneuverability? Journal of Experimental Biology, 203, 3391-3396.
Wassersug, R. J. & Sperry, D. G. (1977). The relationship of locomotion to differental predation on Pseudacris triseriata (Anura: Hylidae). Ecology, 58, 830-839.
Weinstein, R. B. & Full, R. J. (1999). Intermittent locomotion increases endurance in a gecko. Physiological and Biochemical Zoology, 72, 732-739.
Wilson, A. & Lichtwark, G. (2011). The anatomical arrangement of muscle and tendon enhances limb versatility and locomotor performance. Philosophical Transactions of the Royal Society B, 366, 1540-1553.
Woakes, A. J. & Foster, W. A. (eds.) (1991). The comparative physiology of exercise. Journal of Experimental Biology, 160, 1-340.
Ydenberg, R. C. & Dill, L. M. (1986). The economics of fleeing from predators. In Rosenblatt, J. S., Beer, C., Busnel, M.-C. & Slater, P. J. B. (eds.) Advances in the Study of Behavior, Vol. 16. pp. 229-249.
Zaaf, A., Herrel, A., Aerts, P. & De Vree, F. (1999). Morphology and morphometrics of the appendicular musculature in geckoes with different locomotor habits (Lepidosauria). Zoomorphology, 119, 9-22.
Zamparo, P., Perini, R., Orizio, C., Sacher, M. & Ferretti, G. (1992). The energy cost of walking or running on sand. European Journal of Applied Physiology, 65, 183-187.
Zani, P. A. (2000). The comparative evolution of lizard claw and toe morphology and clinging performance. Journal of Evolutionary Biology, 13, 316-325.
Zehr, E. P. & Sale, D. G. (1994). Ballistic movement: muscle activation and neuromuscular adaptation. Canadian Journal of Applied Physiology, 19, 363-378.