ESCAPING FROM PREDATORS An Integrative View of Escape Decisions (2015)
Part III Related behaviors and other factors influencing escape
10 Vigilance, alarm calling, pursuit deterrence, and predation inspection
Guy Beauchamp
10.1 Introduction
Prey species that detect predators late in the predation sequence have little option but to flee as soon as possible. However, prey species may exercise some control over the timing of predator detection, which can free up time to evaluate the threat that the predator poses and even alter the outcome of the encounter with the predator (Caro 2005). The purpose of this chapter is to examine factors that influence predator detection and the options available to a prey animal between the detection of a predator and escape.
The ability to detect predators early in the predation sequence depends on the amount of time allocated to antipredator vigilance. Maintaining a vigilant state allows prey animals to detect predators earlier but at a cost. Indeed, vigilance often conflicts with other fitness-enhancing activities, such as foraging and sleeping, and, generally, prey species must make a trade-off between their various time demands (Caraco 1979). Any factor that influences how much time individuals can allocate to antipredator vigilance will have an impact on the distance at which a predator can be detected on average. I shall review some of the major factors that affect vigilance, such as group size, and highlight some recent developments in vigilance research with a direct bearing on escape behavior. In particular, vigilance may be expected to fluctuate through time within the same group, implying that the ability to detect predators may also vary on a short time scale.
Prey species can manipulate the outcome of an encounter with a predator after detection. Many prey species produce alarm calls upon detection of a predator (Bradbury & Vehrencamp 2011). Such signals may be aimed at the predator to indicate detection, but they often transfer information to nearby conspecifics. Recent work has shown that alarm calls can convey information about predator size and distance and even behavior, which may be crucial to receivers in determining the best course of action as the predator approaches (Zuberbühler 2009). Upon hearing an alarm call, individuals may flee immediately or assess the situation before initiating flight. The second part of this chapter focuses on the properties of alarm calls and their effects on conspecifics.
Signals produced after predator detection can also convey information to the predator (Shelley & Blumstein 2005). Prey animals that are likely to escape following detection may signal to the predator that it has been detected. In addition, they can signal their quality to the predator. Predators may use information from these signals to abandon chases that would most likely be futile. The third part of this chapter examines the effects of signals produced by prey on their predators.
Many prey species approach detected predators rather than flee (Pitcher et al. 1986; FitzGibbon 1994). This counterintuitive tactic, which brings the prey closer to a potentially harmful predator, illustrates the notion that prey are willing to take risk to gather information about the motivation of a predator. Predator inspection behavior is common in fish and some mammals, and I close this chapter by evaluating the costs and benefits of this behavior.
10.2 Antipredator vigilance
If prey animals were in predator detection mode at all times, the distance at which a predator would be detected would simply reflect the ability of the prey to extract threat signals from other non-predator related environmental stimuli, or noise. Basic predator detectability would vary among individuals, and from species to species, reflecting sensory processing ability. In birds, for instance, greater visual acuity associated with larger eyes allows earlier detection (Fernández-Juricic et al. 2004). Sensory processing ability would interact with the degree of noise that characterizes the habitat to determine when an approaching predator first becomes detectable to the prey. For example, visual detection may be less effective in cluttered habitats (Devereux et al. 2006) and aural detection may deteriorate in noisier settings (Quinn et al. 2006).
This ideal scenario is unlikely to apply under most circumstances for at least two reasons. First, rival sources of attention are almost always present in the lives of prey animals. Their attention may be diverted to monitor the activity of neighbors (Favreau et al. 2010) or to fulfill other fitness-enhancing needs such as grooming, sleeping or foraging (Caraco 1979). In fact, the ability to detect predators may be severely compromised when performing these activities. For instance, the senses of a sleeping prey animal are at best intermittently available, which may allow predators to approach undetected more easily (Lima et al. 2005). Animals also need to forage, and again the effectiveness of predator detection may be limited when resource exploitation interferes with detection (Lima & Bednekoff 1999; Kaby & Lind 2003). Second, the ability to detect stimuli may, paradoxically, decrease when vigilance for predation threats is maintained for too long a period of time due to habituation or fatigue (Dimond & Lazarus 1974). This physiological response may entice prey animals to reduce to some extent the total amount of time allocated to predator detection so as to maintain a high level of detectability. This discussion emphasizes that the time available to detect predators is most likely limited.
Overall, the distance at which a predator can be first detected will reflect the interplay between sensory processing ability, the amount of environmental noise, and the time that can be devoted to predator detection. All else being equal, animals that invest less time in predator detection may be expected to detect predators later on average. Any factor that influences the time devoted to predator detection, which is typically referred to as vigilance, is thus likely to impact the ability to detect predators early. In the following, I explore some attributes of vigilance and the role of several ecological and physiological variables known to influence its expression in the context of escape behavior.
10.2.1 What is vigilance?
Vigilance is a state of alertness that allows the detection of relevant stimuli. It is practically measured using external expressions of alertness, such as when an animal raises its head to scan the surroundings. Indeed, a posture with the head up was used to define vigilance in the very first model of vigilance (Pulliam 1973). The focus on head position assumes that vigilance is mostly performed visually, but acoustic vigilance may be a common feature in animals and need not be performed head up (Ridgway et al. 2006). In this case, it may be difficult to assess when an animal is vigilant.
Two types of vigilance are generally distinguished: routine or induced vigilance (Blanchard & Fritz 2007). Routine vigilance occurs during spare time while induced vigilance is viewed as a direct reaction to an imminent predation threat. It is the investment in routine vigilance that matters in the context of predator detection. Induced vigilance would occur after a threat is detected and allow animals to weigh their options before escaping.
10.2.2 Can detection of predators occur when non-vigilant?
Vigilance was initially viewed as being entirely incompatible with other activities such as foraging. This simple dichotomy made it very easy to determine the amount of time spent vigilant by an individual. The incompatibility assumption probably reflects the fact that early models of vigilance focused on species of birds feeding with the head down, a posture thought to impair their ability to detect threats. However, recent research with birds, and other species as well, implies that some level of vigilance may be maintained during other activities. Some species of animals can be alert for predators even when asleep (Rattenborg et al. 1999). When foraging, animals with laterally facing eyes can monitor areas directly above them to some extent (Lima & Bednekoff 1999; Fernández-Juricic et al. 2008; Wallace et al. 2013). Many species can also monitor their surroundings while searching or handling food in a head-up posture (Kaby & Lind 2003; Fortin et al. 2004).
The fact that some level of vigilance may be maintained by non-overtly vigilant animals suggests that predator detection is possible regardless of head position. However, it appears that the ability to detect predators is weaker when the head is down. In an influential series of experiments, Lima and Bednekoff aimed to determine the extent to which non-overtly vigilant animals can detect a predator. When head up, dark-eyed juncos (Junco hyemalis) can detect a fast approaching mounted hawk very rapidly. By contrast, birds with their head down detected the hawk less rapidly, suggesting a less than optimal detection response (Lima & Bednekoff 1999; Figure 10.1). In a further experiment with the same species, individuals increased the duration of their scans to compensate for the addition of visual barriers that prevented head-down vigilance (Bednekoff & Lima 2005), implying that these birds value the information they can acquire when non-overtly vigilant. At least in this species, detection of predators is possible when non-overtly vigilant, but appears less than optimal. Whether this is the case in other species remains to be established. Generally, it may not be obvious to determine when a predator has been detected.
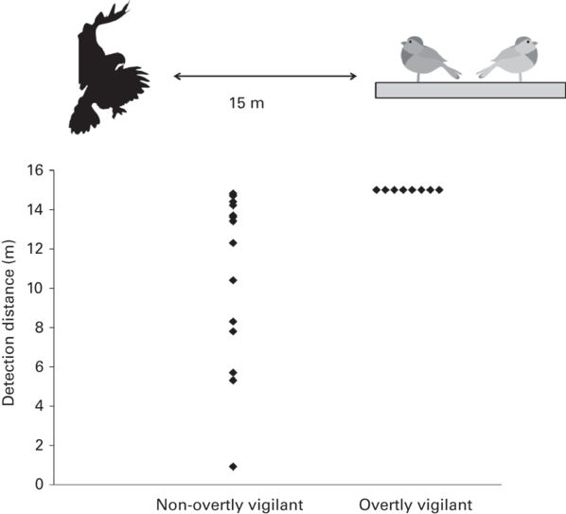
Figure 10.1
Dark-eyed juncos detect a mounted hawk more quickly when overtly vigilant with their head up rather than with their head down. The attack started 15 m away from a feeding platform upon which targeted birds fed.
(After Lima & Bednekoff 1999)
10.2.3 The effect of group size on vigilance
As long as the probability of predator detection is higher during overt vigilance, any factor that reduces time spent vigilant will reduce the ability to detect threats. Vigilance can be influenced by many variables, but the effect of group size has attracted the most attention in the literature (Beauchamp 2008). Models of vigilance predict that individual investment in vigilance should decrease as group size increases for two main reasons (McNamara & Houston 1992). First, as group size increases, more eyes and ears become available to monitor the surroundings so that threats are less likely to go undetected. Once a threat is detected, the fright responses of detectors can warn the non-detectors rapidly, allowing all group members to flee rapidly. Second, the presence of several potential targets in a group effectively dilutes predation risk for each prey as long as the rate of encounter with predators does not increase with group size and predators capture at most one individual per attack. In view of the decreased risk of capture in a group, individuals can reduce their investment in vigilance. As long as the vigilance at the group level is high enough, individuals in larger groups should enjoy greater safety than when foraging alone or in small groups.
Predator detection should thus typically occur sooner in larger groups, which benefit from the presence of many potential detectors. In a classic experiment, Kenward used a trained goshawk (Accipiter gentilis) to attack wood pigeons (Columba palumbus) in groups of varying sizes from a standard distance. As the number of pigeons increased in the group, individuals detected the predator at a greater distance, as predicted from the pooling of individual detection effort (Kenward 1978). While this study only relied on one trained predator, results along these lines have been obtained in other species (Caro 2005).
Detection of threats will not necessarily always occur earlier in larger groups. What matters for detection is the level of vigilance maintained at the group level, which is known as collective vigilance. Detection will occur sooner in larger groups as long as collective vigilance increases with group size. However, as I explained earlier, individuals are expected to be less vigilant in larger groups, implying that collective vigilance is unlikely to increase linearly with group size. Moreover, the time freed by a reduction in vigilance becomes available to increase fitness through other activities, such as foraging. Under some circumstances, the fitness gains that accrue from an increase in foraging may allow individuals to sacrifice safety, leading to a lower level of collective vigilance in large groups. Sacrifices in safety may make sense in various situations, including when foragers have low energy reserves or exploit resources in rich food patches (McNamara & Houston 1992). In such cases, detection ability could, theoretically, be lower in larger groups. Further research should allow us to determine whether the advantages in having more eyes and ears in a larger group always translate into faster detection.
10.2.4 Temporal changes in vigilance
Two recent developments in vigilance research are especially relevant to escape behavior. Collective vigilance is a key factor in determining the ability to detect predators early. These recent developments suggest that collective vigilance may in fact fluctuate through time, which implies that the ability to detect predators may not be constant within the same group. Such fluctuations have been related to two factors: vigilance copying and predation risk assessment.
Temporal fluctuation in collective vigilance is predicted to occur when individuals copy the vigilance of one another (Beauchamp et al. 2012; Sirot & Touzalin 2009). To put matters into perspective, models of vigilance typically assume that vigilance should be independent among group members. Sirot and Touzalin (2009) suggested that individuals in a group should, instead, copy the vigilance of their neighbors under some circumstances detailed below. When copying occurs, collective vigilance will be less than predicted under independent vigilance because there will be more bouts than expected with few or no group members vigilant at the same time.
Why would individuals copy the vigilance of their neighbors if it compromises the detection of predators? As I pointed out earlier, direct detection of threats allows a quicker escape. If predators preferentially target those that escape more slowly, an individual should be more vigilant when its neighbors are vigilant to avoid being left behind when the group is attacked. Copying vigilance is expected to produce rises and falls in the proportion of vigilant group members through time, temporal waves as it were. Copying vigilance has been documented recently in many species of birds and mammals (Beauchamp 2009b; Ge et al. 2011; Michelena & Deneubourg 2011; Öst & Tierala 2011; Pays et al. 2012), where it has been possible to eliminate the possibility that outside stimuli triggered synchronized vigilance. One study also provided evidence for temporal waves of vigilance in sleeping gulls (Beauchamp 2011; Figure 10.2). The implication for escape behavior research is that the ability to detect predators may fluctuate on a short time scale within the same group.
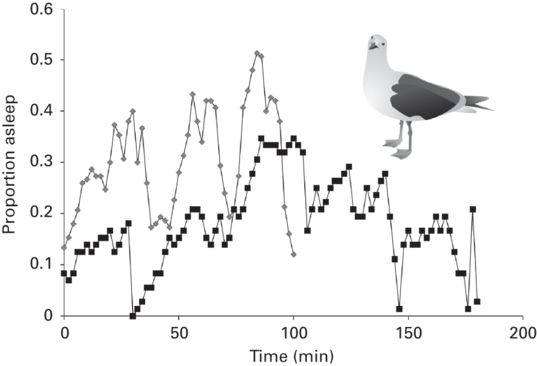
Figure 10.2
Collective temporal waves of sleeping, a low vigilant state, in two roosting flocks of gulls (Larus spp.). Time series analysis revealed a statistically significant periodicity of 46 min in one flock of 72 gulls (dark squares) and of 34 min in a flock of 150 gulls (gray squares).
(After Beauchamp 2011)
Copying vigilance should produce rises and falls in collective vigilance but about the same mean. Recent models suggest that collective vigilance may also be expected to decrease systematically with time since a group started to forage. A decrease in individual vigilance with time may reflect the outcome of an arms race between predators and prey (Beauchamp & Ruxton 2012). Prey species often have little information about current predation risk in the food patches they visit. In particular, a group that arrives at a new patch must determine whether a predator lurks nearby - like a lion (Panthera leo) hiding in the bushes (Périquet et al. 2010). The predator must decide when to break cover, and the prey animals must choose the level of vigilance to adopt as a function of time. If the hiding predator always broke cover at the same time, individuals in the group will be selected to be extra-vigilant until that very time. Predators should thus keep the group guessing about attack time. However, waiting too long may not be desirable as group members may be more likely to detect the predator. Group members should also keep the predator guessing about when vigilance will decrease to avoid the situation in which the predator simply waits for vigilance to come down before launching an attack. The predicted solution to this war of attrition between predator and prey takes the following form: predators attack at unpredictable times but typically early and group members adopt a high vigilance early and then switch unpredictably later to lower vigilance (Beauchamp & Ruxton 2012).
Prey animals in groups do not always face observant predators, but still need to assess predation risk. As time goes by without an attack, individuals may feel safer and progressively and adaptively lower their vigilance (Sirot & Pays 2011). The implication of these directional changes in vigilance is that the ability to detect predators may decrease as foraging progresses in the same group. Lack of control over time since a feeding bout started will thus generally increase the amount of noise in detection distance. In addition, predator detection distance is expected to decrease gradually during a feeding bout. Empirical studies with simulated attacks would be useful to establish whether detection ability and escape behavior change at times where the group is expected to be less vigilant, say, during a trough in collective vigilance or later in a feeding bout.
10.2.5 Other factors that affect vigilance
Many factors other than group size are known to influence the allocation of time to vigilance. All these factors will have an impact on the ability to detect threats by reducing the time devoted to predator detection. Lack of control over these factors can introduce additional variation among samples of detection distance.
Habitat characteristics that influence the ability of escape, time of day, and food availability are three examples of environmental factors that influence vigilance. The ability to escape may vary in certain habitats, due to the presence of obstacles or because protective cover is further away (Lima 1992), for instance, which should influence the level of vigilance that prey animals adopt. Detection ability may thus increase in habitats where the probability of escaping is slighter. Time of day can influence vigilance through many means. Predation risk may vary throughout the day as some predators are more active at particular times of the day or more difficult to detect due to poor light condition (Lima 1988). When predation risk is perceived to be higher, prey animals are expected to increase vigilance, which may allow them to detect predators sooner. Vigilance may also vary on a daily basis in response to changing energy requirements (Pravosudov & Grubb 1998). For instance, vigilance may increase from early morning to the afternoon as animals become more satiated.
Samples of detection distances may often be taken from areas that differ markedly in resource availability and quality. Food patch characteristics can also influence vigilance levels, and thus indirectly detection distances (Beauchamp 2009a). Models indicate that when animals face time constraints or, in other words, when there is a tangible risk of starvation, the level of vigilance should decrease as the rate of food intake increases (McNamara & Houston 1992). As patches of higher quality typically allow foragers to obtain food more rapidly, vigilance is thus expected to decrease with food density. In conclusion, several environmental factors are known to influence vigilance and must be properly controlled, either statistically or experimentally, to reduce the amount of noise in detection distance.
10.3 Alarm calling
Vigilance is an investment in time prior to predator detection. It increases the chances of detecting the predator before it is too late. The following sections focus on what happens after predator detection but prior to the actual escape. After detection, prey animals often emit alarm calls or produce other signals. Here, I am concerned with signals aimed at conspecifics. In the following section, I shall deal with signals aimed at the predator.
In view of the limited amount of time allocated to vigilance by each group member, few prey animals are expected to detect an approaching predator at the same time. Nevertheless, the detection of a threat by a few spreads rapidly throughout the group. Detectors provide indirect cues of detection to non-detectors through their behavior (Lima 1995). For instance, eastern gray kangaroos (Macropus giganteus) that have detected a snake stare intently at the predator and this cue soon alerts the others in the group (Pays et al. 2013). A more active system to alert neighbors is the use of alarm calls, which can reach other group members rapidly even when they are not in direct visual contact. In the context of escape behavior, the occurrence of alarm calls can change the costs and benefits of the options available to group members prior to fleeing, and thus affect the course of action selected.
10.3.1 Alarm calls can convey information about the type of threat
Alarm calls can provide crucial information about the type of threat the group faces and eventually which reaction is the most appropriate. In a classic paper, Seyfarth et al. (1980) showed that different types of predators elicited different alarm calls in free-living vervet monkeys (Chlorocebus pygerythrus). With the use of playbacks, the authors subsequently demonstrated that vervet monkeys adapted their escape tactics to the type of alarm calls broadcasted: climbing in a tree when they heard the playback of an alarm call associated with a leopard (Panthera pardus), looking down when they heard the alarm call elicited by a snake, and running to cover and looking up when they heard the alarm call produced after spotting an eagle (Seyfarth et al. 1980; Figure 10.3). In other species, acoustic features of alarm calls have been shown to convey information about predator size (Templeton et al. 2005), distance to the predator (Wilson & Evans 2012), urgency to respond (Manser et al. 2002), and even predator behavior (Griesser 2008), all of which could allow signal receivers to better choose their escape tactics.
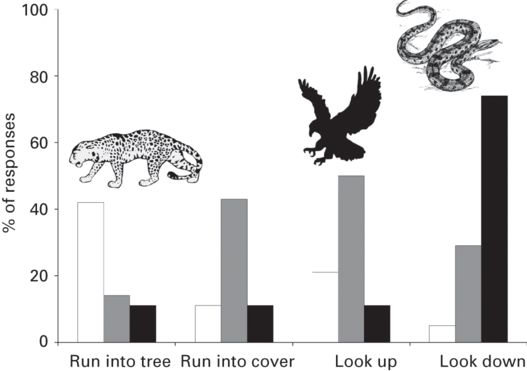
Figure 10.3
Vervet monkeys respond differently to playbacks of alarm calls elicited by a leopard, an eagle, or a snake. The percentage of trials where at least one individual showed a given response for longer after than before the playback is shown for each type of predator (n = 19 for the leopard (white bars), 14 for the eagle (gray bars), and 19 for the snake (black bars)).
(After Seyfarth et al. 1980)
This degree of sophistication in alarm calling probably matches the variety of escape tactics available to a species. A single kind of alarm call may be sufficient to all types of predators when there is only one way to escape (Blumstein & Armitage 1997), but different alarm calls matched to a predator or a specific situation may be beneficial when prey animals can respond to a threat in many different ways, as the vervet example clearly illustrates.
10.3.2 False alarm calls
Alarm calls are notoriously unreliable. Indeed, a high percentage of all alarm calls in many species are actually false alarms attributable to non-threatening stimuli (Cresswell et al. 2000; Blumstein et al. 2004). Upon hearing an alarm call, it would make sense for non-detectors to evaluate the potential threat by themselves rather than flee immediately.
One way to reduce superfluous escapes is to adjust the strength of responses to the number of alarm callers. When many individuals sound the alarm at the same time, it is probably more likely that the threat is real. Supporting this hypothesis, yellow-bellied marmots (Marmota flaviventris) were found to be more vigilant after hearing two alarm callers at the same time rather than one (Blumstein et al. 2004). In birds, flushes to safety by several individuals at the same time are also more effective in triggering responses by nearby companions than single flushes (Lima 1995; Cresswell et al. 2000).
In many species, alarm calls have been reported to be individually distinctive, which raises the possibility that signal receivers may respond differently according to the reliably of alarm callers. Indeed, some individuals may have a higher ratio of false to real alarms due to their lack of experience or a lower threshold for responding to potentially threatening stimuli. This idea was tested by associating the individually distinct calls of some yellow-bellied marmots to a real threat and others to harmless stimuli, varying the reliability of alarm callers (Blumstein et al. 2004). Marmots discriminated among reliable and less reliable callers and spent more time investigating the source of disturbance following broadcasts of calls from the unreliable callers.
10.3.3 Alarm calls and antipredator ploys
Alarm calls may signal an imminent threat to which immediate action is required. However, the threat may not always be urgent, and in this case alarm calling may be useful to deploy antipredator tactics that will reduce the chances of a successful attack. Alarm calls in many species cause signal receivers to adopt cryptic behavior such as crouching, whose purpose appears to be to reduce the likelihood of detection by the predator (Evans et al. 1993). Long-distance alarm calls in one species of monkey, indicative of a distant threat, causes individuals to bunch together, increasing the effectiveness of factors such as collective detection and risk dilution (Shultz et al. 2003). Alarm calls often trigger an increase in vigilance in signal receivers thus increasing the number of individuals actually monitoring the threat (Loughry & McDonough 1988). Increased vigilance may be useful to better assess a threat (perhaps reducing the rate of false alarms) or to ready all group members for mass flight (see below).
10.3.4 Alarm calls and mass flight
By warning other group members rapidly, alarm calls may play yet another function prior to escape. Rather than fleeing alone, an alarm caller can effectively recruit all group members for mass flight (Owens & Goss-Custard 1976; Sherman 1985), potentially reducing the chances of capture through the confusion or selfish-herd effects.
Confusion acts when the predator closes in on a group of prey and attempts to capture a particular individual from a fleeing group. Reduced capture rate after initiating a chase is thought to reflect distraction of attention from the targeted prey by the presence of several non-target companions (Neill & Cullen 1974; Landeau & Terborgh 1986). Recent research suggests that confusion is a line of defense that works by exploiting the cognitive limitations of the predator (Tosh et al. 2006). The empirical evidence that confusion plays a role during escape behavior following an alarm call is rather anecdotal (Leger et al. 1980; Sherman 1985). To be more convincing, we would need data on the effect of prey density on predator success rate with the prediction that capture rate should decrease when prey are closer to one another.
The alarm caller could also benefit by seeking protection inside the group, effectively using other group members as a shield (Hamilton 1971). To invoke the selfish-herd effect, we need direct evidence that alarm callers select particular trajectories in space that would reduce their domain of danger when fleeing. It is interesting to note that if an alarm caller chooses to flee, the range of options for signal receivers becomes very limited. As Sherman (1985) pointed out, a signal receiver that fails to flee cannot benefit from the confusion or selfish-herd effects and may also provide a more inviting target to the predator.
10.4 Pursuit-deterrence signals
I explore signals made by prey animals upon detection of a predator focusing this time on their potential effects on predators. As with alarm calling, these signals can change the value of the choices available to prey after detection. If the predator, say, abandons the chase after receiving the signal, the prey animals may choose to maintain vigilance rather than flee.
The logic of signaling to an approaching predator is simple: when an individual is quite likely to avoid capture, it makes sense for the individual to produce a signal to dissuade the predator from attacking. Capture may be unlikely if, for example, the predator has been detected early and lost the element of surprise or if the prey animal possesses attributes, such as speed, that make capture more challenging. With such a signal, prey can avoid the energy and time costs of an actual escape while the predator avoids the costs of a most likely futile pursuit (Ruxton et al. 2004). Such signals have been referred to as pursuit-deterrent signals. Pursuit-deterrence signals come in two types: signals that advertise detection to the predator and those that signal the condition of the signaler to the predator.
10.4.1 Perception advertisement
This type of signal may be produced in two contexts. The most obvious one is when the predator is detected outside the typical range of attack distances. Beyond this range, the risk of capture is low and producing the signal does not reduce the ability to escape should the predator choose to attack. Other means of dissuading the predator from attacking after detection include mobbing by the prey (Tan et al. 2012) or hiding in a refuge, both of which significantly reduce the chances of capture. Examples of detection signals involve visual signals such as tail flagging in deer (Caro et al. 1995) and ground squirrels (Barbour & Clark 2012), tail wagging in lizards (Cooper 2010), or auditory signals such as long-distance calling in primates (Zuberbühler et al. 1997).
Perception advertisement signals may also be produced when the prey animals strongly suspect that a predator is present (Bergstrom & Lachmann 2001). This mechanism involves parsing information from the environment and producing the signal when a threshold of perceived risk has been crossed. These perception advertisement signals may thus be made in the absence of a predator.
The costs of a perception advertisement signal for the prey involve the production of the signal and the risk of inadvertently attracting the attention of other predators. Producing the signal itself is probably inexpensive, certainly in comparison to avoiding the costs of a chase altogether. Signaling, especially by mistake, can be costly if it attracts the attention of undetected predators. This potential cost may not apply when predator density is low and when the risk of mistaking harmless stimuli as real is also low. From the predator point of view, the benefits include the probability of capture during a chase and the costs include the time and energy involved in the chase. Mathematical modeling reveals that a perception advertisement signal can be evolutionarily stable if the benefits of producing the signals exceed the costs for the signaler and if the costs of chasing are high and the probability of capture is low for the predator (Bergstrom & Lachmann 2001).
While it is clear that both predator and prey can benefit from perception advertisement signals, it is less clear what prevents prey from cheating. Indeed, prey animals could produce signals of detection more frequently than needed in the odd chance that a predator has approached undetected. However, this is not a stable solution in view of the time costs involved in the production of needless signals and the chances that the signal may attract the attention of other predators, thus increasing predation risk inadvertently. Nevertheless, pre-emptive perception signals may work if signals have a short range, which makes them unlikely to attract other predators, and if the signal is typically associated with honest detection (Murphy 2007).
10.4.2 Quality advertisement
If the odds of capture after detecting the predator are high, it would seem at first ill-advised for the prey to take the time to produce a pursuit-deterrent signal when it could flee instead and gain more ground on the predator. Individuals that have the ability to escape at closer distances may still benefit from producing such signals, effectively advertising their quality to the predator. To evolve, such signals must be costly otherwise all individuals, those able to escape or not, will produce the signal making it useless (Grafen 1990). Honest signals may include those that are condition dependent. For example, the length or frequency or intensity of a signal may be indicative of the condition of the signaler, which would allow the predator to judge the quality of the prey. Mathematical modeling reveals that such signals can evolve as long as there is a close relationship between features of the signal and condition of the prey, and that the risk of capture for signaling prey decreases as the condition of the signaler increases (Vega-Redondo & Hasson 1993). These signals are considered quality advertisement signals, and tend to be more costly than the perception advertisement signals discussed above (Caro 2005).
What prevents cheating in this signaling system is the relationship between specific features of the signal and individual quality. A low-quality individual is simply unable to produce signals that would deceive the predator (Zahavi & Zahavi 1997). Intriguingly, cheating may be preventable even if the relationship between signal features and individual quality is rather weak. Consider a situation in which low- and high-quality individuals produce the same signal. If a low-quality individual finds itself in a group with n - 1 high-quality companions, the low-quality individual will still have a 1/n chance of being targeted by a predator that attacks any group member with the same probability. The risk of capture for the low-quality individual is higher now because the predator has been allowed much closer during the display. In addition, the poor quality of the prey animal will hamper its escape. Cheating of this type may only be possible in very large groups in which the risk of being targeted is much lower.
While predators should give up quite readily after a perception advertisement signal, there should be a close relationship between abandoning a hunt and some definable features of a quality advertisement signal. The difficulty is to distinguish between the two types of signals. It is not always obvious to determine the costs of a signal, and, moreover, the quality of the prey may not be easy to assess in the field. Stotting is a good case in point. When stotting, an animal jumps up in the air with all four legs held stiffly. The success rate of attacks by cheetahs (Acynonyx jubatus) on Thomson’s gazelles (Gazella thomsoni) decreases from 21 to 0% when the quarry stots (Caro 1986). Stotting may signal to the predator that it has been detected, but may also signal the quality of the signaler. The crucial data on the relationship between stotting performance and condition of the prey species come from a study involving African wild dogs (Lacyon pictus), another predator of Thomson’s gazelles (FitzGibbon & Fanshawe 1988). Indeed, African wild dogs were more likely to chase gazelles that stotted at a lower rate (Figure 10.4). After being targeted, gazelles that eventually escaped stotted longer during the chase than those that were captured (Figure 10.4). Supporting the hypothesis that stotting reveals the condition of the prey, stotting was more common during the wet season when body condition generally improves. In other species, stotting rate correlates positively with body condition (Caro 1994).
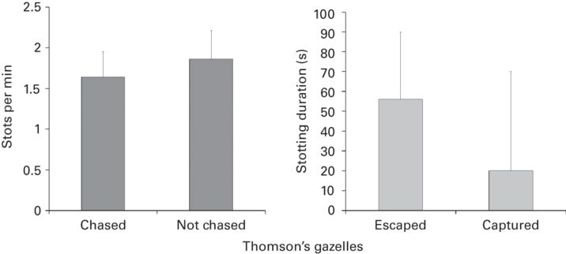
Figure 10.4
African wild dogs prefer to chase Thomson’s gazelles that stotted at a lower rate on average (left panel). Error bars show one standard deviation. Gazelles that stotted longer during the chase were less likely to be captured (right panel). The median and the 75th percentile are illustrated.
(After FitzGibbon & Fanshawe 1988)
10.4.3 Evolution of pursuit-deterrent signals
Signals produced by a solitary animal after the detection of a predator, such as an alarm call, can only be aimed at the predator. By contrast, signals produced by animals living in groups may be aimed at companions as well as the predator. To distinguish between the two potential targets of such signals, it is necessary to understand the context of signaling. If group-living animals produce such signals when they are alone, the predator is certainly a potential target (Murphy 2006) although this does not rule out that the signal may also be useful to warn companions.
As signals that evolved in one context may be co-opted by another, the current target of a signal may not reflect very well the selection pressures that led to its evolution in the first place. A good example of this comes from the evolution of white tails in rails, a clade of reclusive semi-aquatic birds (Stang & McRae 2009). Rails flick their tails when disturbed and this behavior is more common in species with white tails, which contrast sharply with their overall dark plumage. This display may be used to signal to the predator that it has been detected and/or to alert companions in social species. A phylogenetic reconstruction showed that rails living in open habitats are more likely to have white tails. In addition, the transition to open habitats tended to precede the evolution of white tails while gregariousness evolved later in some lineages. This ordering of events indicates that white tails probably evolved as an antipredator signal needed in the more risky open habitats, and any current use by conspecifics reflects a co-option of the signal for other purposes.
10.5 Predator inspection
After detecting a predator, several options are available to prey animals in groups. When an attack is imminent, the only option is to flee as soon as possible. However, predators are not always in attack mode and, in this case, it may pay foragers to assess the risk posed by the predator before initiating flight. Predator inspection behavior appears to play just this purpose. During predator inspection, a subset of individuals leave the group, approach the predator, stop, and eventually return to the group. This behavior is best known in schooling species of fish and mammalian herbivores (Magurran 1986; FitzGibbon 1994; Caro et al. 2004). Predator inspection probably provides information about the location and motivation of the predator, which can be passed along to companions in the group. Predator inspection may be helpful to identify situations where the predator is uninterested in attack, which would allow prey to save the cost of increased vigilance and fleeing needlessly.
10.5.1 Why inspect predators?
At first sight, predator inspection appears ill-advised. Why risk death by approaching a potentially dangerous predator rather than simply hide in the safety of the group? Early research showed that inspectors do assess the risk associated with approaching a predator. In one fish species, predator inspection was indeed more likely in less threatening situations (e.g., immobile predator) and when many individuals joined the inspection party (Pitcher et al.1986). Fish also tend to avoid the cone of attack of a predator during inspection by biasing their approach toward the tail rather than the head (Brown & Dreier 2002). Similarly, Thomson’s gazelles are more likely to approach a predator when in large groups and in a terrain that reduces the probability of ambush. Nevertheless, predator inspection is riskier than remaining behind in the group (FitzGibbon 1994; Milinski et al. 1997), suggesting that predator inspection must provide compensating benefits.
One possibility is that predator inspection can actually reduce predation risk for the inspectors. Indeed, guppies (Poecilia reticulata) that inspected a predator were less likely to be attacked and killed than those that remained behind (Godin & Davis 1995; Figure 10.5). Similar findings have been reported in European minnows (Phoxinus phoxinus; Magurran 1990) and Thomson’s gazelles (FitzGibbon 1994).
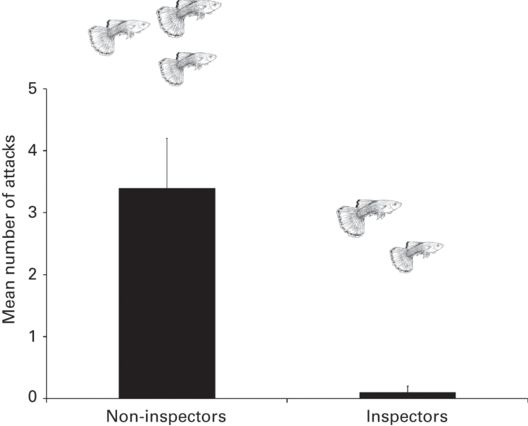
Figure 10.5
Guppies that inspect a fish predator are attacked less often than those that remain behind. Error bars show one standard deviation.
(After Godin & Davis 1995)
Predator inspection can reduce predation risk through many means. The approach of several inspectors may confuse the predator, which may then prefer to attack those that remain behind (Curio 1978). Inspection may signal to the predator that it has been detected and that an attack on them rather than on those left behind is most likely futile.
Supporting the idea that inspection reduces the probability of attack, cheetahs tended to move to a different area after having been approached by inspecting Thomson’s gazelles. Finally, inspection may also be a signal of quality to discourage the predator from attacking those that comes closest. Support for the quality advertisement hypothesis comes from the observation that larger three-spined sticklebacks (Gasterosteus aculeatus) and those in better condition are more likely to approach predators (Külling & Milinski 1992).
Predator inspection is often performed in groups and appears relatively contagious. In Thomson’s gazelles, for example, all members of a group typically approach the predator and other groups nearby are also likely to join (FitzGibbon 1994). Approaching in groups certainly reduces the probability of attack for any individual group member through a simple dilution of risk. Risk dilution reduces the costs of predator inspection.
10.5.2 Preferences for partners
In the preceding section, I showed that individuals can discriminate among alarm callers on the basis of their reliability. Research on predator inspection behavior has also examined whether the choices of partners during approaches to a predator are similarly biased. As predator inspection involves risks, an individual that remains slightly behind during predator inspection would gain all the benefits from the inspection without incurring the risks. To avoid such cheating, it was proposed that individuals should seek partners that have proven reliable during previous predator inspection bouts, a tit-for-tat like behavior (Milinski 1987). Evidence from the laboratory and the field suggests that fish can form strong ties with one another while foraging and during predator inspection, which suggests that reciprocity may be involved in choosing partners for inspection (Croft et al. 2006). However, crucial evidence that individuals are less likely to carry out predator inspection with partners that have defected in the past is generally lacking (Thomas et al. 2008). These issues suggest at the very least that group composition may influence the occurrence of predator inspection.
10.6 Future directions
The amount of time between predator detection and escape is under the control of the prey animals to some extent and can be increased by allocating more time to antipredator vigilance. This period allows individuals to assess the threat posed by a predator and to communicate through signals with nearby conspecifics and the predator. The risks of being attacked and of escaping can be manipulated by using signals after predator detection. In the following, I highlight some specific challenges for future work on the topics covered in this chapter.
Antipredator vigilance represents an option to increase the distance at which predators are detected on average. It is not an easy task to determine detection distance since vigilance serves many purposes and varies as a function of several environmental variables. A greater understanding of the factors that influence collective vigilance will be needed to make more precise predictions. Recent work also suggests that different individuals may vary consistently in their vigilance (Carter et al. 2009; Mathot et al. 2011). This finding fits with the observation that animals can vary in the level of risk they are prepared to accept (Reale et al. 2007). This raises the fascinating possibility that detection at the group level may vary as a function of the phenotypic composition of the group, and that different individuals may be able to detect predators more quickly than others. Future work along these lines will probably challenge the assumption that all individuals adopt the same vigilance in a group.
The type of information conveyed by alarm calls has been studied intensely, but we still know little about the costs and benefits of uttering alarm calls. It is usually thought that such signals reduce the personal safety of the callers, but the evidence is limited at the moment (Caro 2005). Many of the mechanisms that may allow prey animals to escape following an alarm, such as the confusion and selfish-herd effects, remain to be demonstrated convincingly. Complicating matters, alarm calls may not only warn conspecifics but also act concomitantly as a signal to the predator, making it difficult to disentangle the respective selection pressures. The information from alarm calls can also be used by nearby individuals from different species to mount effective antipredator responses (Goodale et al. 2010), effectively increasing the audience network for such calls. It will be interesting to determine in the future what information members of different species can extract from alarm calls developed for members of one species, and whether such calls can influence the detection ability and escape behavior of other species.
By contrast to other means of dissuading predators from attacking, such as crypsis or aposematic signals, pursuit-deterrent signals are displayed on a facultative basis, which highlights the costs of using such signals at all times and also makes it possible to show graded signals that are condition dependent. It would be interesting to investigate why a facultative tactic serves the purposes of predator deterrence to a greater extent in some species.
Predator inspection behavior can also be viewed as a pursuit-deterrent signal. If it serves as a detection signal, it is not clear why detection cannot be signaled from the confine of the group at a safe distance from the predator. Coming closer to the predator is risky, but in fish at least inspection behavior deflects the costs to those behind. This situation would make sense if inspection behavior is a quality advertisement signal. However, the costs and benefits of predator inspection behavior are known to some extent in a relatively small number of species and general conclusions are difficult to draw.
References
Barbour, M. A. & Clark, R. W. (2012). Ground squirrel tail-flag displays alter both predatory strike and ambush site selection behaviours of rattlesnakes. Proceedings of the Royal Society B: Biological Sciences, 279, 3827-3833.
Beauchamp, G. (2008). What is the magnitude of the group-size effect on vigilance? Behavioral Ecology, 19, 1361-1368.
Beauchamp, G. (2009a). How does food density influence vigilance in birds and mammals? Animal Behaviour, 78, 223-231.
Beauchamp, G. (2009b). Sleeping gulls monitor the vigilance behaviour of their neighbours. Biology Letters, 5, 9-11.
Beauchamp, G. (2011). Collective waves of sleep in gulls (Larus spp.). Ethology, 117, 326-331.
Beauchamp, G., Alexander, P. & Jovani, R. (2012). Consistent waves of collective vigilance in groups using public information about predation risk. Behavioral Ecology, 23, 368-374.
Beauchamp, G. & Ruxton, G. D. (2012). Changes in anti-predator vigilance over time caused by a war of attrition between predator and prey. Behavioral Ecology, 23, 368-374.
Bednekoff, P. A. & Lima, S. L. (2005). Testing for peripheral vigilance: do birds value what they see when not overtly vigilant? Animal Behaviour, 69, 1165-1171.
Bergstrom, C. T. & Lachmann, M. (2001). Alarm calls as costly signals of antipredator vigilance: The watchful babbler game. Animal Behaviour, 61, 535-543.
Blanchard, P. & Fritz, H. (2007). Induced or routine vigilance while foraging. Oikos, 116, 1603-1608.
Blumstein, D. T. & Armitage, K. B. (1997). Does sociality drive the evolution of communicative complexity: A comparative test with ground-dwelling sciurid alarm calls. American Naturalist, 150, 179-200.
Blumstein, D. T., Verneyre, L. & Daniel, J. C. (2004). Reliability and the adaptive utility of discrimination among alarm callers. Proceedings of the Royal Society of London - Series B: Biological Sciences, 271, 1851-1857.
Bradbury, J. W. & Vehrencamp, S. L. (2011). Principles of Animal Communication, 2nd edn. Sunderland: Sinauer.
Brown, G. E. & Dreier, V. M. (2002). Predator inspection behaviour and attack cone avoidance in a characin fish: The effects of predator diet and prey experience. Animal Behaviour, 63, 1175-1181.
Caraco, T. (1979). Time budgeting and group size: A theory. Ecology, 60, 611-617.
Caro, T. M. (1986). The functions of stotting in Thomson’s gazelles: Some tests of the predictions. Animal Behaviour, 34, 663-684.
Caro, T. M. (1994). Ungulate antipredator behaviour: Preliminary and comparative data from African bovids. Behaviour, 128, 189-228.
Caro, T. M. (2005). Antipredator Defenses in Birds and Mammals. Chicago, IL: University of Chicago Press.
Caro, T. M., Lombardo, L., Goldizen, A. W. & Kelly, M. (1995). Tail-flagging and other antipredator signals in white-tailed deer: New data and synthesis. Behavioral Ecology, 6, 442-450.
Caro, T. M., Graham, C. M., Stoner, C. J. & Vargas, J. K. (2004). Adaptive significance of antipredator behaviour in artiodactyls. Animal Behaviour, 67, 205-228.
Carter, A. J., Pays, O. & Goldizen, A. W. (2009). Individual variation in the relationship between vigilance and group size in eastern grey kangaroos. Behavioral Ecology & Sociobiology, 64, 237-245.
Cooper, W. E. (2010). Timing during predatorprey encounters, duration and directedness of a putative pursuit-deterrent signal by the zebra-tailed lizard, Callisaurus draconoides. Behaviour, 147, 1675-1691.
Cresswell, W., Hilton, G. M. & Ruxton, G. D. (2000). Evidence for a rule governing the avoidance of superfluous escape flights. Proceedings of the Royal Society B: Biological Sciences, 267, 733-737.
Croft, D., James, R., Thomas, P. et al. (2006). Social structure and co-operative interactions in a wild population of guppies (Poecilia reticulata). Behavioral Ecology and Sociobiology, 59, 644-650.
Curio, E. (1978). The adaptive significance of avian mobbing: I. Teleonomic hypothesess and predictions. Zeitscrift fur Tierpsychologie, 48, 175-183.
Devereux, C. L., Whittingham, M. J., Fernández-Juricic, E., Vickery, J. A. & Krebs, J. R. (2006). Predator detection and avoidance by starlings under differing scenarios of predation risk. Behavioral Ecology, 17, 303-309.
Dimond, S. & Lazarus, J. (1974). The problem of vigilance in animal life. Brain, Behavior and Evolution, 9, 60-79.
Evans, C. S., Evans, L. & Marler, P. (1993). On the meaning of alarm calls: Functional reference in an avian vocal system. Animal Behaviour, 46, 23-38.
Favreau, F.-R., Goldizen, A. W. & Pays, O. (2010). Interactions among social monitoring, anti-predator vigilance and group size in eastern grey kangaroos. Proceedings of the Royal Society B: Biological Sciences, 277, 2089-2095.
Fernández-Juricic, E., Erichsen, J. T. & Kacelnik, A. (2004). Visual perception and social foraging in birds. Trends in Ecology & Evolution, 19, 25-31.
Fernández-Juricic, E., Gall, M. D., Dolan, T., Tisdale, V. & Martin, G. R. (2008). The visual fields of two ground-foraging birds, House Finches and House Sparrows, allow for simultaneous foraging and anti-predator vigilance. Ibis, 150, 779-787.
FitzGibbon, C. D. (1994). The costs and benefits of predator inspection behaviour in Thomson’s gazelles. Behavioral Ecology & Sociobiology, 34, 139-148.
FitzGibbon, C. D. & Fanshawe, J. (1988). Stotting in Thomson’s gazelles: An honest signal of condition. Behavioral Ecology & Sociobiology, 23, 69-74.
Fortin, D., Boyce, M. S., Merrill, E. H. & Fryxell, J. M. (2004). Foraging costs of vigilance in large mammalian herbivores. Oikos, 107, 172-180.
Ge, C., Beauchamp, G. & Li, Z. (2011). Coordination and synchronisation of anti-predation vigilance in two crane species. Plos One, 6, e26447.
Godin, J.-G. J. & Davis, S. A. (1995). Who dares, benefits: Predator approach behaviour in the guppy (Poecilia reticulata) deters predator pursuit. Proceedings of the Royal Society of London. Series B: Biological Sciences, 259, 193-200.
Goodale, E., Beauchamp, G., Magrath, R. D., Nieh, J. C. & Ruxton, G. D. (2010). Interspecific information transfer influences animal community structure. Trends in Ecology & Evolution, 25, 354-361.
Grafen, A. (1990). Biological signals as handicaps. Journal of Theoretical Biology, 144, 517-546.
Griesser, M. (2008). Referential calls signal predator behaviour in a group living bird. Current Biology, 18, 69-73.
Hamilton, W. D. (1971). Geometry for the selfish herd. Journal of Theoretical Biology, 31, 295-311.
Kaby, U. & Lind, J. (2003). What limits predator detection in blue tits (Parus caeruleus): Posture, task or orientation? Behavioral Ecology & Sociobiology, 54, 534-538.
Kenward, R. E. (1978). Hawks and doves: Factors affecting success and selection in goshawk attacks on woodpigeons. Journal of Animal Ecology, 47, 449-460.
Külling, D. & Milinski, M. (1992). Size-dependent predation risk and partner quality in predator inspection of sticklebacks. Animal Behaviour, 44, 949-955.
Landeau, L. & Terborgh, J. (1986). Oddity and the “confusion effect” in predation. Animal Behaviour, 34, 1372-1380.
Leger, D. W., Owings, D. H. & Gelfand, D. L. (1980). Single-note vocalizations of California ground squirrels: Graded signals and situation-specificity of predator and socially evoked calls. Zeitscrift fur Tierpsychologie, 52, 227-246.
Lima, S. L. (1988). Vigilance during the initiation of daily feeding in dark-eyed juncos. Oikos, 53, 12-16.
Lima, S. L. (1992). Strong preferences for apparently dangerous habitats? A consequence of differential escape from predators. Oikos, 64, 597-600.
Lima, S. L. (1995). Collective detection of predatory attack by social foragers: Fraught with ambiguity? Animal Behaviour, 50, 1097-1108.
Lima, S. L. & Bednekoff, P. A. (1999). Back to the basics of antipredatory vigilance: can nonvigilant animals detect attack? Animal Behaviour, 58, 537-543.
Lima, S. L., Rattenborg, N. C., Lesku, J. A. & Amlaner, C. J. (2005). Sleeping under the risk of predation. Animal Behaviour, 70, 723-736.
Loughry, W. J. & McDonough, C. M. (1988). Calling and vigilance in California ground squirrels: A test of the tonic communication hypothesis. Animal Behaviour, 36, 1533-1540.
Magurran, A. E. (1986). Predator inspection behaviour in minnow shoals: Differences between populations and individuals. Behavioral Ecology & Sociobiology, 19, 267-273.
Magurran, A. E. (1990). The adaptive significance of schooling as antipredator defence in fish. Annals Zoologi Fennici, 27, 51-66.
Manser, M. B., Seyfarth, R. M. & Cheney, D. L.(2002). Suricate alarm calls signal predator class and urgency. Trends in Cognitive Sciences, 6, 55-57.
Mathot, K. J., van den Hout, P. J., Piersma, T. et al. (2011). Disentangling the roles of frequency vs. state-dependence in generating individual differences in behavioural plasticity. Ecology Letters, 14, 1254-1262.
McNamara, J. M. & Houston, A. I. (1992). Evolutionarily stable levels of vigilance as a function of group size. Animal Behaviour, 43, 641-658.
Michelena, P. & Deneubourg, J.-L. (2011). How group size affects vigilance dynamics and time allocation patterns: The key role of imitation and tempo. Plos One, 6, e18631.
Milinski, M. (1987). Tit-for-tat in sticklebacks and the evolution of cooperation. Nature, 325, 433-437.
Milinski, M., Lüthi, J. H., Eggler, R. & Parker, G. A. (1997). Cooperation under predation risk: Experiments on costs and benefits. Proceedings of the Royal Society of London. Series B: Biological Sciences, 264, 831-837.
Murphy, T. G. (2006). Predator-elicited visual signal: Why the turquoise-browed motmot wag-displays its racketed tail. Behavioral Ecology, 17, 547-553.
Murphy, T. G. (2007). Dishonest “preemptive” pursuit-deterrent signal? Why the turquoise-browed motmot wags its tail before feeding nestlings. Animal Behaviour, 73, 965-970.
Neill, S. R. S. J. & Cullen, J. M. (1974). Experiments on whether schooling by their prey affects the hunting behaviour of cephalopods and fish predators. Journal of Zoology, 172, 549-569.
Öst, M. & Tierala, T. (2011). Synchronized vigilance while feeding in common eider brood-rearing coalitions. Behavioral Ecology, 22, 378-384.
Owens, N. W. & Goss-Custard, J. D. (1976). The adaptive significance of alarm calls given by shorebirds on their winter feeding grounds. Evolution, 30, 397-398.
Pays, O., Beauchamp, G., Carter, A. J. & Goldizen, A. W. (2013). Foraging in groups allows collective predator detection in a mammal species without alarm calls. Behavioral Ecology, 24, 1229-1236.
Pays, O., Sirot, E. & Fritz, H. (2012). Collective vigilance in the Greater Kudu: Towards a better understanding of synchronization patterns. Ethology, 118, 1-9.
Périquet, S., Valeix, M., Loveridge, A. J. et al. (2010). Individual vigilance of African herbivores while drinking: the role of immediate predation risk and context. Animal Behaviour, 79, 665-671.
Pitcher, T. J., Green, D. A. & Magurran, A. E. (1986). Dicing with death: Predator inspection behaviour in minnow shoals. Journal of Fish Biology, 28, 439-448.
Pravosudov, V. V. & Grubb, T. C.(1998). Body mass, ambient temperature, time of day, and vigilance in tufted titmice. Auk, 115, 221-223.
Pulliam, H. R. (1973). On the advantages of flocking. Journal of Theoretical Biology, 38, 419-422.
Quinn, J. L., Whittingham, M. J., Butler, S. J. & Cresswell, W. (2006). Noise, predation risk compensation and vigilance in the chaffinch Fringilla coelebs. Journal of Avian Biology, 37, 601-608.
Rattenborg, N. C., Lima, S. L. & Amlaner, C. J. (1999). Half-awake to the risk of predation. Nature, 397, 397-398.
Reale, D., Reader, S. M., Sol, D., McDougall, P. T. & Dingemanse, N. J. (2007). Integrating animal temperament within ecology and evolution. Biological Reviews, 82, 291-318.
Ridgway, S., Carder, D., Finneran, J. et al. (2006). Dolphin continuous auditory vigilance for five days. Journal of Experimental Biology, 209, 3621-3628.
Ruxton, G. D., Sherratt, T. N. & Speed, M. P. (2004). Avoiding Attack: The Evolutionary Ecology of Crypsis, Warning Signals and Mimicry. Oxford: Oxford University Press.
Seyfarth, R. M., Cheney, D. L. & Marler, P. (1980). Vervet monkey alarm calls: Semantic communication in a free-ranging primate. Animal Behaviour, 28, 1070-1094.
Shelley, E. L. & Blumstein, D. T. (2005). The evolution of vocal alarm communication in rodents. Behavioral Ecology, 16, 169-177.
Sherman, P. W. (1985). Alarm calls of Belding’s ground squirrels to aerial predators: Nepotism or self-preservation? Behavioral Ecology and Sociobiology, 17, 313-323.
Shultz, S., Faurie, C. & Noë, R. (2003). Behavioural responses of Diana monkeys to male long-distance calls: Changes in ranging, association patterns and activity. Behavioral Ecology and Sociobiology, 53, 238-245.
Sirot, E. & Pays, O. (2011). On the dynamics of predation risk perception for a vigilant forager. Journal of theoretical Biology, 276, 1-7.
Sirot, E. & Touzalin, F. (2009). Coordination and synchronization of vigilance in groups of prey: The role of collective detection and predators’ preference for stragglers. American Naturalist, 173, 47-59.
Stang, A. T. & McRae, S. B. (2009). Why some rails have white tails: The evolution of white undertail plumage and anti-predator signaling. Evolutionary Ecology, 23, 943-961.
Tan, K., Wang, Z., Li, H. et al. (2012). An “I see you” prey-predator signal between the Asian honeybee, Apis cerana, and the hornet, Vespa velutina. Animal Behaviour, 83, 879-882.
Templeton, C. N., Greene, E. & Davis, K. (2005). Allometry of alarm calls: Black-capped chickadees encode information about predator size. Science, 308, 1934-1937.
Thomas, P. O. R., Croft, D. P., Morrell, L. J. et al. (2008). Does defection during predator inspection affect social structure in wild shoals of guppies? Animal Behaviour, 75, 43-53.
Tosh, C. R., Jackson, A. L. & Ruxton, G. D. (2006). The confusion effect in predatory neural networks. American Naturalist, 167, E52-E65.
Vega-Redondo, F. & Hasson, O. (1993). A game-theoretic model of predator-prey signaling. Journal of Theoretical Biology, 162, 309-319.
Wallace, D. J., Greenberg, D. S., Sawinski, J. et al. (2013). Rats maintain an overhead binocular field at the expense of constant fusion. Nature, 498, 65-69.
Wilson, D. R. & Evans, C. S. (2012). Fowl communicate the size, speed and proximity of avian stimuli through graded structure in referential alarm calls. Animal Behaviour, 83, 535-544.
Zahavi, A. & Zahavi, A. (1997). The Handicap Principle: A Missing Piece of Darwin’s Puzzle. Oxford: Oxford University Press.
Zuberbühler, K. (2009). Survivor signals: The biology and psychology of animal alarm calling. Advances in the Study of Behavior, 40, 277-322.
Zuberbühler, K., Noë, R. & Seyfarth, R. M. (1997). Diana monkey long-distance calls: Messages for conspecifics and predators. Animal Behaviour, 53, 589-604.