BioBuilder: Synthetic Biology in the Lab (2015)
Chapter 10. Golden Bread
BioBuilder’s Golden Bread activity emphasizes the “build” phase of the design-build-test cycle. You will work with a strain of baker’s yeast that has been modified with genes from another fungus to produce β-carotene, a nutrient we naturally obtain from eating foods such as carrots, sweet potatoes, and broccoli. In the body, β-carotene is converted to vitamin A, which is crucial for vision, the immune system, and other biological functions. In some developing countries that struggle with malnutrition, vitamin A deficiency is a critical public-health issue. Researchers hope that an engineered strain of baker’s yeast designed to generate β-carotene, like the one you will investigate in this activity, could be used in bread to treat vitamin A deficiency. Such bread might appear a golden color from the added vitamin, hence the name, “golden bread.”
For any new food or drug to become widely available for use, manufacturers must show that they can reliably produce the material. It can’t vary dramatically between batches. It must be consistently effective. In fact, reliability is crucial for nearly all engineering endeavors. If you think about your favorite engineered object, whether it’s your car, your cell phone, or your refrigerator, the fact that it behaves reliably is probably part of what makes it a favorite. People hate when their car won’t start, when their phone reception is poor, and when their refrigerator warms up. We hold these objects to pretty high standards of performance. In fact, some engineered things like bridges, ATMs, and voting machines must work perfectly all the time. So, it’s fair to say that, to be useful, all engineered systems should be reliable.

In the field of synthetic biology, reliability engineers identify sources of instability in a living system to make the system operate more dependably. They have their work cut out for them! BioBuilder’s Golden Bread activity emphasizes how a scientific approach can be combined with good old engineering know-how to evaluate and then improve an unreliable synthetic cell.
Engineering Reliability
If the civil engineers in your hometown have done their job, you don’t find driving over a bridge to be a terrifying experience. You don’t hold your breath the entire way across, worried that the structure will buckle under your weight or crash to the ground. Now that you think about it, though, where does this confidence come from? How do we know that bridges are safe?
The answer is that bridges and other construction projects have been engineered for reliability. The designers know that bridge failures could be fatal, expensive, and crippling to a city’s infrastructure, so they make every effort to design and construct a reliable system. In this chapter, we introduce some specific approaches to reliable system design, such as performing routine scheduled maintenance and building with materials that are stronger than minimally necessary. We also discuss how these principles can be extended to the engineering of biological systems.
Scheduled Maintenance
Imagine how great it would be if your car never needed a tune-up. You’d never have to take it to the shop, and it would always perform as perfectly as the day it came off the assembly line. You’d save tons of time and money, and you’d never be inconvenienced by a breakdown. Most drivers would probably be pretty happy to own a car that ran 100 percent reliably.

But drivers and car manufacturers, even if they dream of such a perfect future, know there’s no such thing as a completely reliable car. Cars endure physical wear and tear from friction on the moving parts of the engine, oil that becomes dirty, and tires that need more or less air depending on the season. Engineers have done what they can to maximize the reliability of each component in a working car, but it’s unreasonable to expect all the parts to function perfectly indefinitely. Instead, manufacturers balance a robust system that supports normal use of the car with a simple maintenance plan to keep the car functioning well for as long as possible.
So what’s an engineer to do, knowing that, over time, systems are bound to corrode and then fail? In most manufacturing fields, engineers will determine a mean time to failure, or MTF, for the system and its components. This determination helps designers predict when a system will break. It also guides the designer on when and how to intervene through regular maintenance of the system. Engineers include MTF calculations in their design process so that they can recommend when parts should be serviced and how to use them for greatest longevity.
In one of engineering professor Henry Petroski’s wonderful books, To Engineer is Human, he explores the MTF for a familiar object, namely a paperclip. As you probably know from experience, if you bend a paperclip enough times, it will eventually break, as is demonstrated in Figure 10-1. From a physical perspective, this break occurs because bending puts stress on the metal, which adds physical defects to the material and as the defects accumulate, the metal breaks. You can measure the MTF for a paperclip by counting the number of bends you can introduce before it breaks. To gain confidence in your number, you would repeat the experiment a few times and under a few different conditions to determine an average MTF. Of course, most of us are not worried about repairing paperclips because they are so easily replaced, but imagine that you had a very valuable paperclip. In that case you would keep track of the number of bends that it was exposed to so that you would know when you were nearing the MTF. It’s hard to imagine what “maintenance” could be done on a paperclip, but if that paperclip were valuable enough, there would probably be something to try.
When applying these ideas to biology, it’s clear that the cell has most of its maintenance routine built in. Biological systems are much more dynamic than mechanical structures such as cars or paperclips. Most cells are in a near constant state of turnover as they grow and then pass their genetic instructions to new cells. Cellular growth and division reliably lead to a supply of fresh cells. Cells also repair themselves, at least when the damage to them isn’t too extreme. These aspects of biology, which are discussed in the Fundamentals of Synthetic Biology chapter, are some of what makes synthetic biology so attractive. Synthetic biologists have a dynamic material to build with (cells!) and so must make good use of biology’s features and abilities.
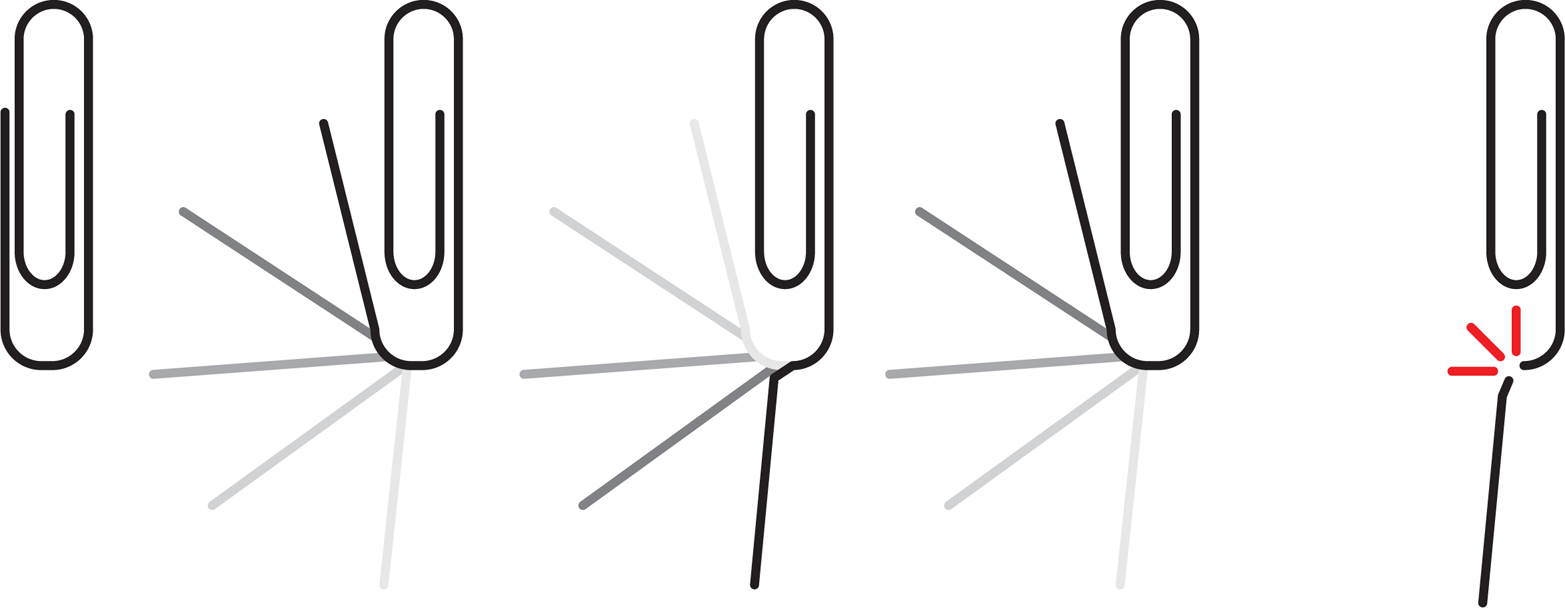
Figure 10-1. A paperclip’s MTF. Bending a paperclip back and forth, as shown here, will eventually cause it to break. The number of bends before breaking can be used to calculate MTF.
Cells’ innate ability to perform routine maintenance also has a downside. Each time a cell makes a new copy of itself to replace a worn out version, the process of cell division introduces mutations into the DNA that are passed to the daughter cells. As a result there is generation-to-generation variation. Such variation can affect the cell’s behavior, and has led to the clever and beautiful systems we find in nature. The variation also presents a significant hurdle for synthetic biologists. The mutations that affect the behavior of the system can take over the population, fully replacing the original cells with in new cells that no longer carry out the designed function (Figure 10-2).
Thus, to engineer a reliable living system, synthetic biologists must address unintended genetic changes that arise over time. As we’ve seen in other BioBuilder chapters, the parallels between more established engineering disciplines and synthetic biology—imperfect as they are—can be helpful, and next we consider some other approaches to engineering reliability.
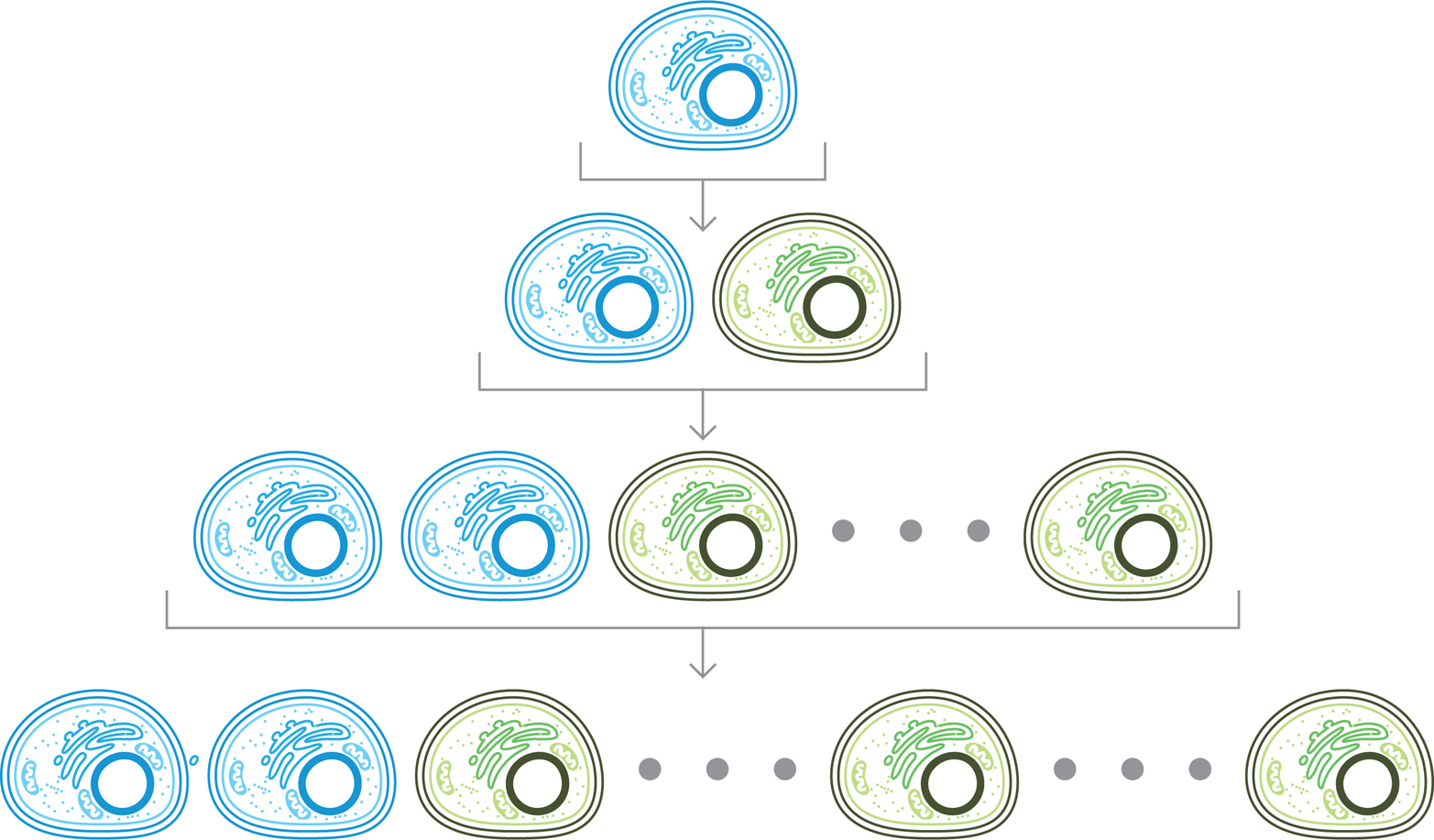
Figure 10-2. Generational variability. Division of an engineered cell (top, blue) can give rise to an exact copy (second row, blue) or a mutated variant (second row, green) that might not carry out the desired function. If the variant is more robust, it can take over the population after a few rounds of division, as shown in the bottom row.
Redundancy
Building redundancy into an engineered system is another technique used in many fields to ensure more reliable performance. For example, engineers working on passenger safety have chosen to deploy both seat belts and air bags in a car crash. The seat belt and airbag systems work independently to meet the same goal. Their independent control makes it less likely that both will fail in a crash. They are also differentially effective. Depending on the angle of the crash, the seat belts or the airbags might have a greater role in protecting the people in the car. Having two safety systems increases the likelihood that passengers will be unharmed in a crash, and so they are worth the added cost. However, not all resource-intensive redundancies are justifiable (Figure 10-3). Consider instead a less life-threatening engineering question: designing a mobile phone. Consumers generally expect their mobile phones to last for only a few years. Screens can crack, software will need updates, and physical features on the newest phones improve. Because most consumers replace their phones rather than live with many of these shortcomings that arise over time, the phone’s designers aren’t motivated to engineer much redundancy into the phone’s electrical components. A failure of the electrical system, perhaps caused by the phone becoming wet or dropped on a hard surface, is inconvenient but not life threatening. It can be remedied by replacing the phone and consumers would rather replace their phones often or buy insurance than increase retail prices for the phones themselves.

Figure 10-3. Redundancy. Wearing both suspenders and a belt is a classic example of designed redundancy.
So, redundancy is unevenly applied across other engineering disciplines and is a resource-intensive approach to reliability, yet it turns out to be a relatively common feature in natural biological systems. For example, most animal cells carry two complete copies of their genome, and plants go even further, sometimes carrying six or eight copies of their genome. The redundancy in living cells is important for their survival, because DNA can be damaged by mutagens in the cell’s environment. Exposure to sunlight or mutagens can induce changes in the DNA sequence, making some of the genetic instructions essentially unreadable. Genetic redundancy provides the cell with some insurance. Even if one of the DNA copies is damaged, another will likely remain intact and can also serve as a template for repairing the damaged copy. The cell must pay a substantial price for this insurance in terms of energy and raw materials, but the cost for the “extra” copies of the genome is repaid in the security of its genetic instructions. Given that genetic redundancy is a standard feature of most multicellular organisms, the evolutionary advantages must outweigh the costs.
Synthetic biologists can introduce genetic redundancy by engineering their system with multiple copies of particular DNA sequences. This can be done by transforming the cells with additional plasmids or by inserting extra copies of certain genes into the cell’s genome. These genetic tricks can enhance the stability of a gene circuit by providing the cell with a backup copy of a gene to use if its other copy fails. Surprisingly, though, it can also destabilize the system. Some organisms, such as yeast, have learned to recognize and remove direct repeats of DNA. There is also a danger of expressing too much of a gene product. Cells are exquisitely efficient systems, so maintaining and expressing extra genetic material can be too energetically costly, adding a metabolic burden to the cell or throwing off the balance in the engineered system. There are ways to work around these pitfalls, such as codon shuffling a redundant gene as we do in the Golden Bread experiment, or adding some regulation to the expression of a redundant gene. The cost/benefit analysis, though, isn’t easy, and it’s still unclear what general rules should be followed when engineering reliability into living systems.
Building Robust Systems
The final approach to engineering reliability that we will discuss here is building a system that is more robust than strictly necessary. As an example, when building a bridge, the engineers could design with materials that would support 60-ton trucks, knowing that the maximum legal weight for trucks in most states is 40 tons. Similarly they could design their bridge to withstand an earthquake measuring 10 on the Richter scale, knowing that the largest earthquake ever recorded is 9.5. To adequately design such robust systems, engineers can use modeling methods that anticipate how their design and materials will react to trucks and earthquakes. The Picture This chapter has more information about modeling and considers how models can provide good data for systems under different stresses.
A disadvantage of designing overly robust systems is the associated cost. Materials to build such a super-strong bridge, for example, might be much more expensive than the ones capable of withstanding the actual expected stresses for the bridge. Most product designers choose materials rationally, often comparing the strength of possible materials to their cost by using a tool called an Ashby Chart. Often, they also factor in the additional time associated with engineering overly robust systems and the cost of that time. The extra-fancy materials might be more complex to build with, to source, or to maintain than the standard ones built in a sufficient system. Another of Henry Petroski’s wonderful books, The Evolution of Useful Things, dedicates much of its chapter “Big Bucks from Small Change” to describing a seemingly mundane engineering project, namely the engineering of slats on a bedframe. Pertoski traces the question of why the slats run square with the frame and not diagonally across it (Figure 10-4). It turns out that this seemingly straightforward question has been posed as far back as Aristotle and is considered in Homer’s The Odyssey when Odysseus builds a bridal bed that must be attached to an olive tree and threaded with leather straps. Surprisingly, the economy and maintenance of a bed with diagonal slats has been deemed unjustifiable for most of recorded history!

Figure 10-4. Balancing robustness and cost. A bed with just horizontal slats (left) is not as strong as a bed with a diagonal grid of slats (right), but the former is less expensive than the latter. Over the years, engineers have repeatedly opted for the less expensive design in this case.
What does material selection look like when it’s extended to the design of biological systems? Perhaps “strength” refers to the expression levels of different proteins in the system, to the predictable growth of the chassis itself, or to the physical properties of the proteins used for the parts and devices. The relevant aspects of the biological system that need to be strengthened will depend on the specific design and its intended usage.
Consider a system like the arsenic sensor from the Fundamentals of Biodesign chapter, or any other environmental sensor for that matter. The biological analog to using super-strong steel for a bridge could be designing the system with many, many copies of the protein sensor device or using a sensor that is more sensitive than strictly necessary. With these modifications, you can rest assured that even a trace amount of the triggering compound will be detected by a sensor and start the cellular reactions you intend. Also, the sensor can continue to function even as deleterious mutations in the gene accumulate over time, because it is already designed to function beyond specifications. The trade-off in this case is the potential for triggering false alarms. The high-affinity arsenic sensor might now recognize similar, less-toxic compounds or even turn ON in the absence of any ligand at all. A system that codes for such high production of the sensor protein might also be metabolically “expensive,” taking lots of the cell’s energy to maintain the high production levels. Many of the cell’s resources will be spent solely on the designed system, as opposed to the other baseline metabolism and repair systems that are needed for cells to remain healthy. Using cellular resources to engineer reliability can end up negatively affecting the health of the cell and so interfere with the engineered system’s function. In fact, the metabolic stress could slow the growth of the cells and decrease the performance of the engineered system—the exact opposite of what the engineer was trying to do when introducing more copies of the sensor.
In addition, if the engineered cells are at an evolutionary disadvantage, they will be replaced by new mutants that grow faster, as discussed previously. Depending on the growth rate of the engineered cells and their mutated versions, the evolutionary battle can be lost in just a few generations. Not surprisingly, then, synthetic biologists are deploying genetic design principles to make their parts “stronger” in the sense that they are less likely to be mutated. For example, because identical DNA sequences are often deleted through recombination, direct repeats of genetic elements are generally avoided. Synthetic biologists can vary the sequence of redundant DNA parts using degeneracies in the genetic code, encoding a leucine with either TTG or CTA, for instance. This trick makes it possible for the same protein to be encoded by two distinct DNA sequences that aren’t so prone to recombine. And by encoding the function with two genes, it might make the system behave more reliably. You’ll directly test this notion with the Golden Bread engineering activity.
Synthetic biologists also have some tools to enhance the chassis they use, making them less vulnerable to unintended changes (Figure 10-5). There are strains available that have improved DNA repair mechanisms. Others have been modified to diminish mutation-inducing processes or to enrich machinery that helps proteins fold appropriately, which may allow a slightly mutated protein to function properly. Finally, some chassis are designed with a “kill switch,” or a means to self-destruct, so any cells that mutate away from the desired behavior can be removed from the population. None of these are failsafe, however. Consequently, one of synthetic biology’s active research areas is the design of safer, more reliable chassis.
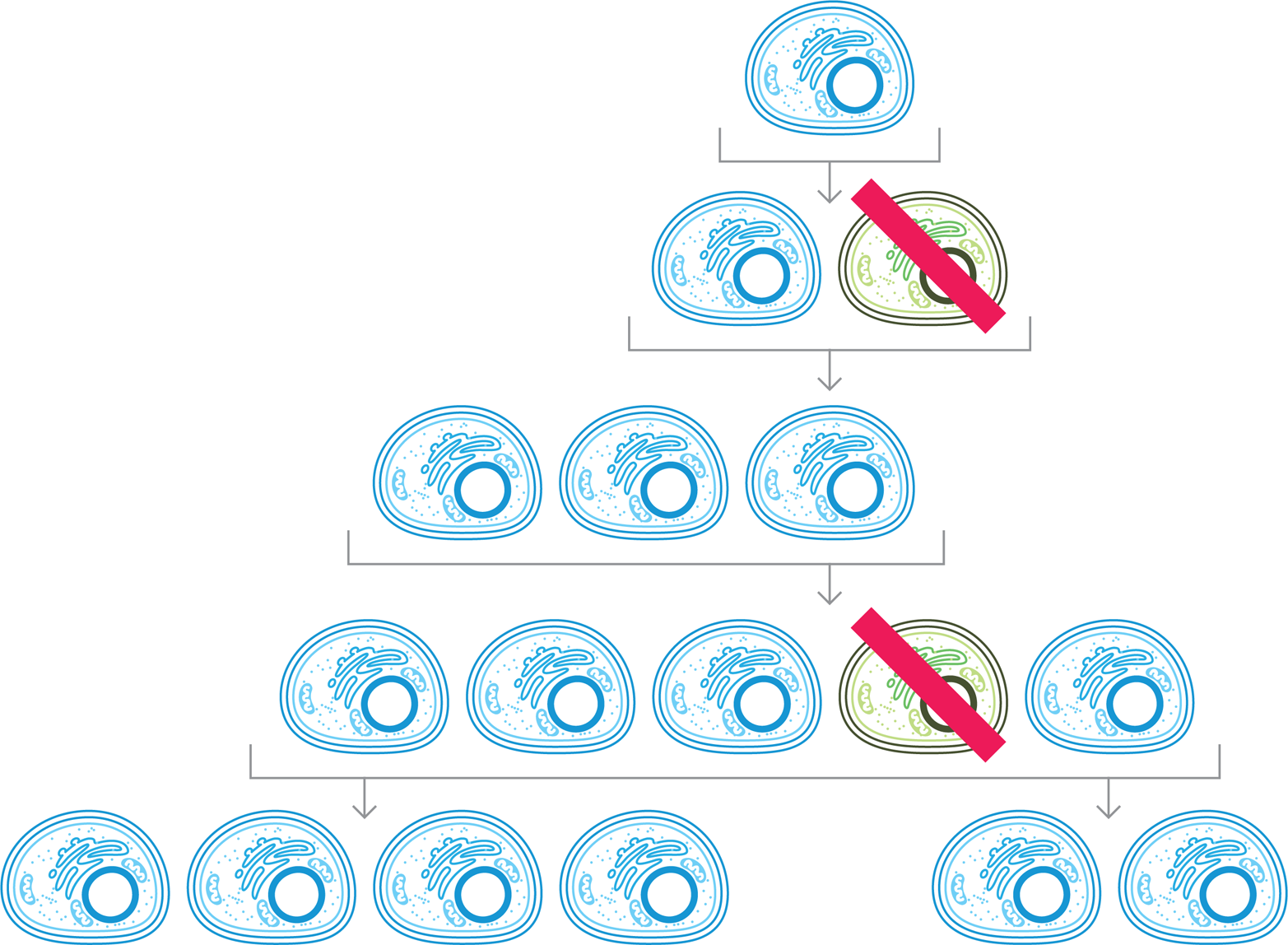
Figure 10-5. A self-correcting system. Division of an engineered cell (top, blue) can create an exact copy (second row, blue) or it can lead to the production of a genetic variant (second row, green) that may not carry out the desired function. To keep these undesired variants from persisting in the population, engineers can design components to kill these cells, as indicated by the red line, to ensure that the system functions reliably.
Background on the “VitaYeast” iGEM Project
Motivation
The only way to get enough vitamin A is through a healthy diet or vitamin supplements. Over 200 million preschool-age children worldwide can’t get vitamin A by either means, and their vitamin A deficiencies can cause significant impact on their vision and their overall health. Vitamin A, which is processed in the body from β-carotene, is a precursor for retinal, a chemical needed for vision. It is also a precursor for retinoic acid, which is crucial for healthy immune systems and development. Nutritional concentrations of vitamin A are found in sweet potatoes, carrots, broccoli, and leafy greens, but vitamin A deficiency (VAD) is a major health problem in places where these foods don’t grow or where they aren’t routinely consumed.
The World Health Organization estimated that vitamin A supplements could reduce deaths of children less than 5 years old by 24 to 34 percent. However, distributing supplements to rural areas can be difficult and costly, and so is not a sustainable solution to this health challenge. Consequently, some researchers aimed to develop a staple food that is itself enriched in vitamin A. In 2000, a European research team announced that it had developed such a food: a strain of “Golden Rice” that was genetically manipulated to express β-carotene. The rice was modified to express three enzymes that would make β-carotene from a natural compound in rice called geranylgeranyl diphosphate. The presence of β-carotene, which is also known as “pro-vitamin A” because it can be converted to vitamin A in the body (Figure 10-6), made the rice grains appear a golden-yellow color, hence the name “Golden Rice” (Figure 10-7).
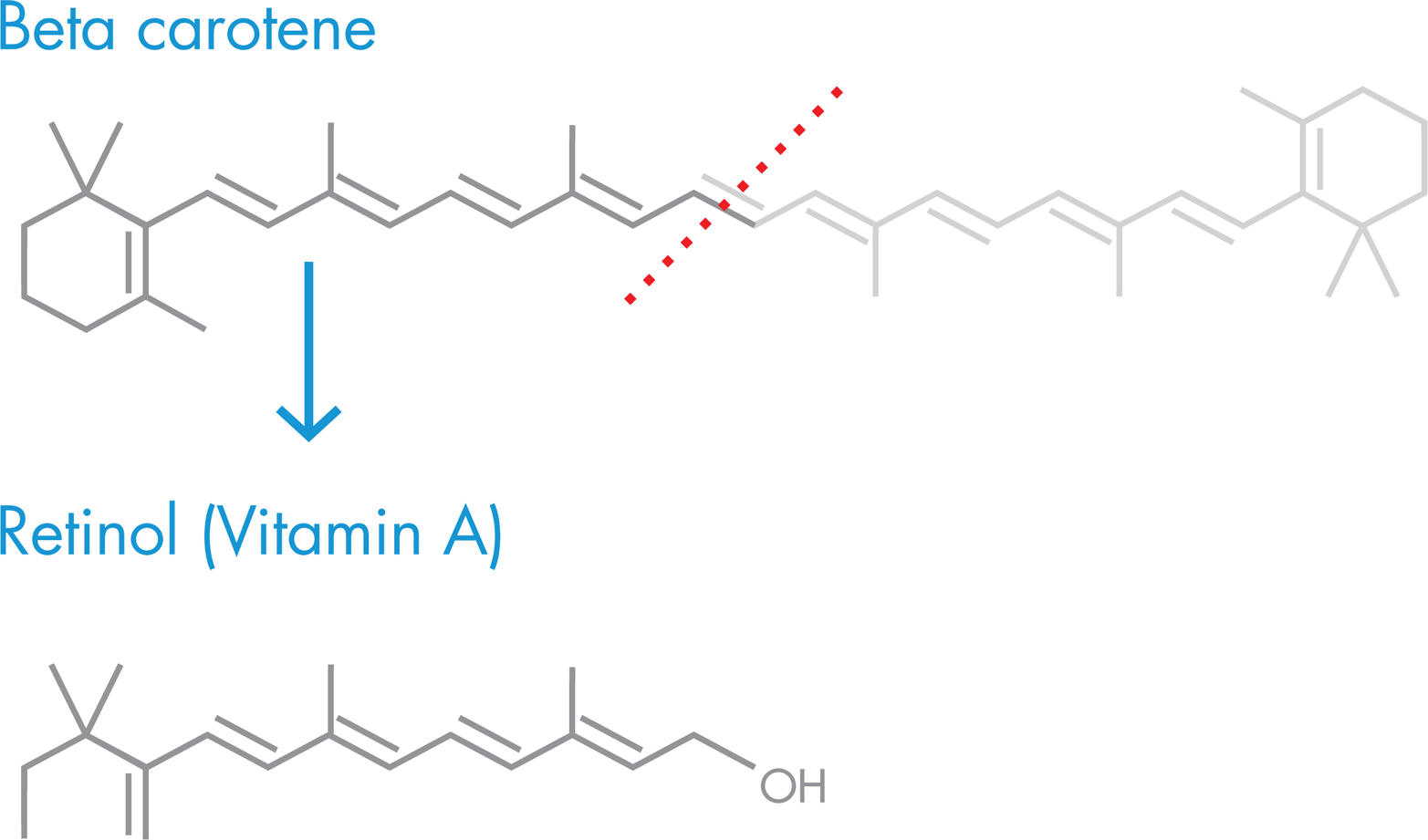
Figure 10-6. Vitamin A synthesis. Humans acquire vitamin A, also known as retinol, by eating a diet rich in β-carotene, which is cleaved into two molecules of vitamin A during digestion.
The researchers hoped to distribute the enriched rice in places where rice is a major component of the local diet and where VAD is a health challenge. However, they encountered significant opposition. Environmental groups and individuals are concerned about potential negative effects of bioengineered food on the health of individuals in the region, on the ecosystem, and on the economy. The opposition to Golden Rice has restricted its distribution, leaving the question of whether it can reduce death from VAD unanswered. The beliefs and conceptions that underlie many of these concerns can initiate a vibrant classroom discussion, and the Fundamentals of Bioethics chapter provides approaches to leading such discussions.
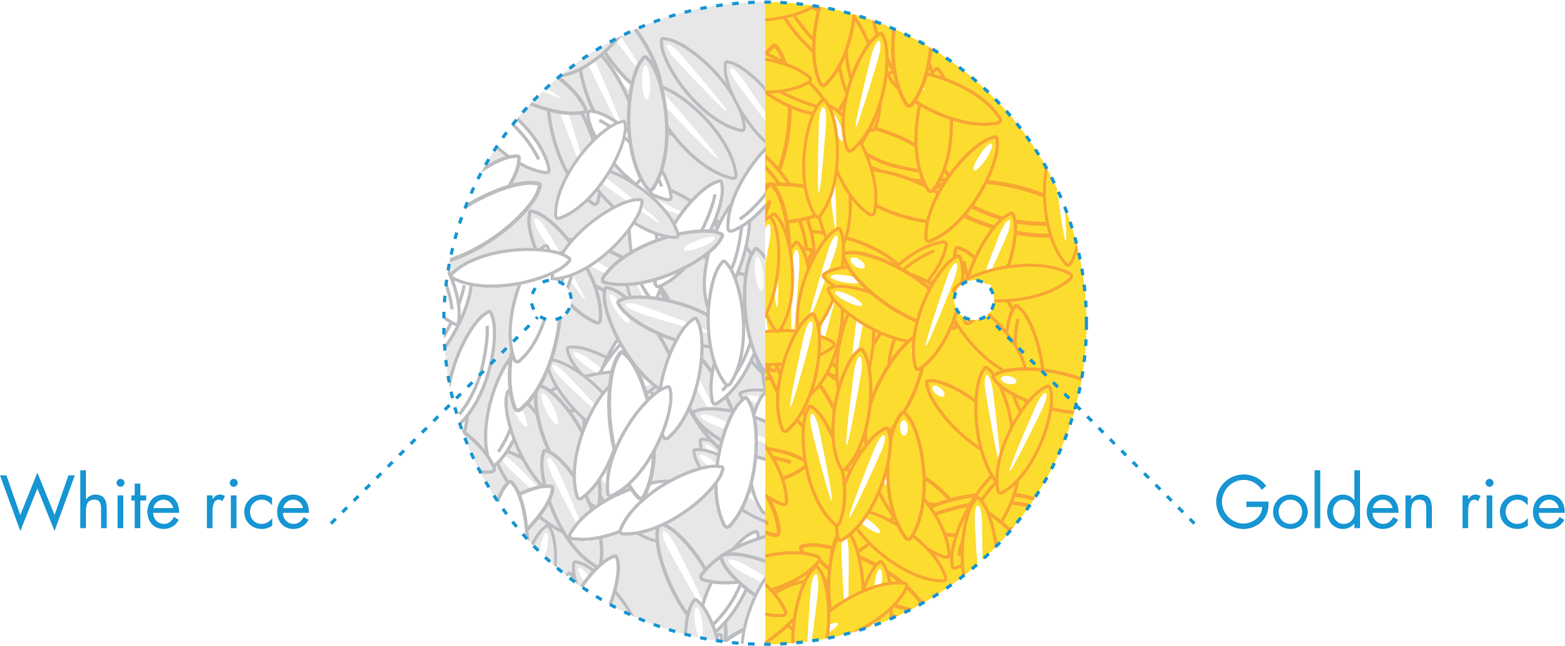
Figure 10-7. Golden rice. Golden rice is white rice that has been enriched to produce β-carotene, which is converted to vitamin A upon ingestion.
The 2011 International Genetically Engineered Machines (iGEM) team from Johns Hopkins University sought a different solution to the VAD/Golden Rice impasse. Rather than add β-carotene to a major food source and change its color to something unnatural, the team decided to work with an engineered version of baker’s yeast, extending some work published in 2007 by researchers who genetically manipulated a common strain, Saccharomyces cerevisiae, to express three β-carotene biosynthesis genes in addition to their normal complement of genetic information (Figure 10-8). The idea the iGEM team had was for this yeast to substitute for standard baker’s yeast, making it possible for users to bake nutrient-enriched loaves of bread. No special baking instructions would be required. Users would simply add a bit of the engineered “VitaYeast” to their standard bread recipe (Figure 10-9). The iGEM team hoped that this solution would alleviate some of the concerns that arose around Golden Rice. The engineered yeast would be a minor component of the vitamin-fortified bread. In addition, the yeast would be killed in the baking of the bread, and the resulting loaves should appear a natural color rather than synthetic and unappetizing.
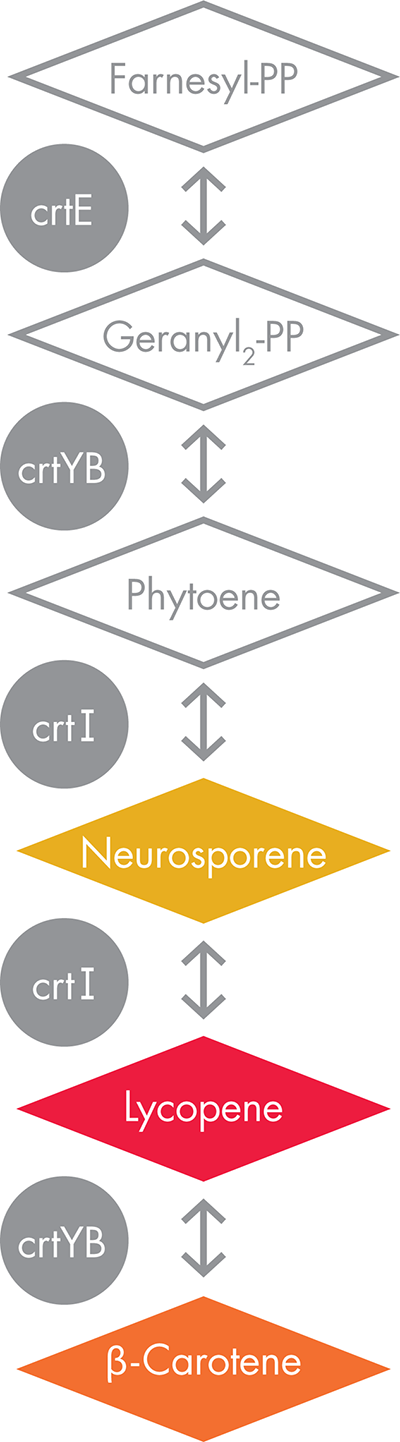
Figure 10-8. A β-carotene biosynthetic pathway. The metabolic pathway used for engineering VitaYeast consists of three enzymes (grey circles) that convert farnesyl phosphate (top diamond) to β-carotene (bottom diamond). The first three compounds in this pathway are colorless, and the last three are colored yellow, red, and orange, respectively. The enzyme function shown here as crtE is naturally present in baker’s yeast but is encoded by a gene called BTS1; the other two enzymes, crtYB and crtI, are imported as genes from a red fungus, X. dendrorhous.

Figure 10-9. Enriched yeast. One day, a commercially available “VitaYeast” could be used instead of baker’s yeast to address vitamin A deficiencies around the globe.
Parts-Level and Device-Level Design
Although the iGEM team’s “VitaYeast” project was motivated by the Golden Rice project and has many similarities, the biology of the rice and the yeast systems have important differences. For this reason (and possibly others), the genetic circuits in the VitaYeast were sourced from a fungus, whereas the genetic circuits for the Golden Rice were sourced from a plant. The researchers who first developed the VitaYeast strain introduced a biosynthetic pathway from a red-colored yeast, Xanthophyllomyces dendrorhous, which naturally produces a carotenoid compound and which has enzymes that can be harnessed for β-carotene production. By contrast, the Golden Rice developers introduced three plant genes to convert geranylgeranyl diphosphate, which is naturally present in rice, to β-carotene.
Like rice, the baker’s yeast S. cerevisiae can naturally produce farnesyl diphosphate, a starting compound for β-carotene synthesis. These yeast, however, also express an enzyme, encoded by the BTS1 gene, that converts the farnesyl diphosphate to geranylgeranyl diphosphate. Converting geranylgeranyl diphosphate into β-carotene requires the action of two enzymes, crtYB and crtI, which were imported into baker’s yeast from the red-colored yeast. Each of these enzymes serves double duty. The crtYB enzyme plays a role early in the synthesis, converting geranylgeranyl diphosphate into phytoene and then comes back into play for the last step of the synthesis, converting lycopene into β-carotene. Between the crtYB-catalyzed steps are two reactions that require the activity of the crtI enzyme, which was also imported into the baker’s yeast strain from the red-colored fungus. The enzyme converts the phytoene first to neurosporene and then to lycopene.
Nature has provided a simple way to detect pigment produced by this pathway. Yeast making β-carotene turn bright orange (Figure 10-10). Alternatively, yeast only making lycopene turn red like a tomato, which has a naturally high lycopene concentration. Finally, neurosporene is a yellow intermediate in this pathway. The color-coding stops there, though, because the earlier compounds in the pathway have fewer than seven conjugated double bonds and are therefore colorless. Nevertheless, the three products in this pathway that are colored made it easier to determine which enzymes work and which strains make the desired compound.
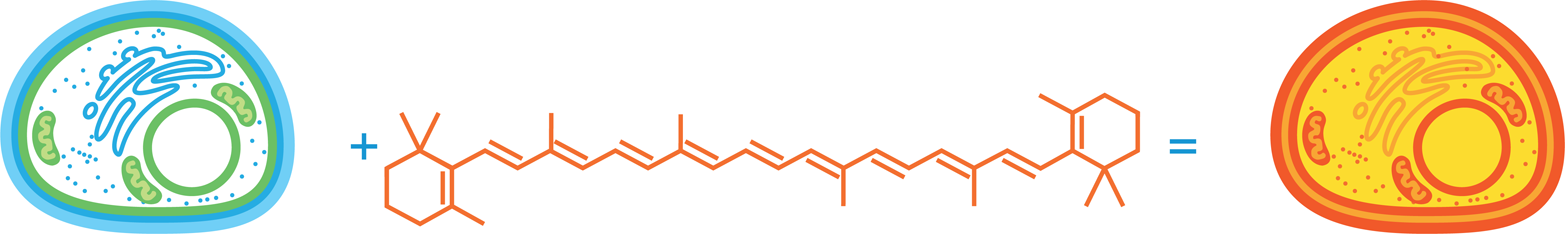
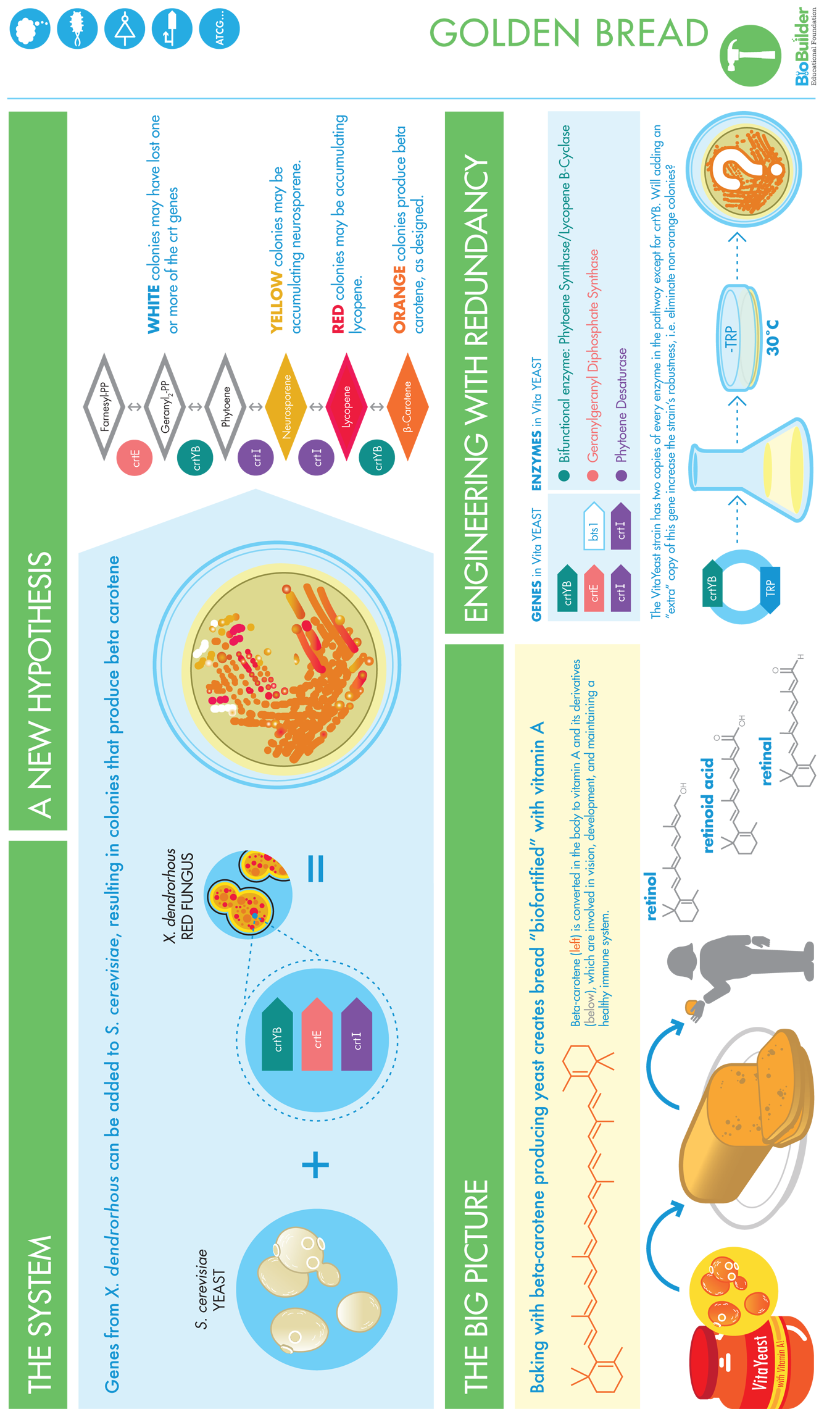
Figure 10-10. VitaYeast design. Engineering baker’s yeast (left) by adding an enzymatic pathway that synthesizes β-carotene (middle) results in a novel, orange-colored, β-carotene synthesizing yeast that can be used for baking.
The researchers who constructed the VitaYeast strain were probably ecstatic when they saw its orange color. Looking closely at what they’d made, however, they must have quickly realized that not all the colonies were behaving as they expected and hoped. Despite the fact that they had all come from the same genetic parent and should have been genetically identical themselves, some were orange, others were red, some were yellow, and some were white. One possible hypothesis is that these differently colored yeast were “stuck” at an intermediate step in the biosynthetic pathway. Understanding whether this hypothesis is in fact the cause of the multicolored yeast and fixing the problem is at the heart of BioBuilder’s Golden Bread activity.
The researchers who designed and built the initial “VitaYeast” strain took two approaches to improving the reliability of its β-carotene production. First, they tried to build a more robust system. In particular, they stopped using the easy-to-work-with plasmids that expressed the genes from the red-colored yeast. Instead of working with plasmids, they moved the crtYB and crtI genes into the chromosome of the baker’s yeast they were building. These integrated copies of the genes are less likely to be lost, and so the strains should be more reliably orange colored. The disadvantage to this approach is that the genes are also harder to work with when they are in the chromosomes because the genetic material can’t be as easily manipulated in the lab, but in this case the researchers felt the improved reliability of the system would be worth the increased trouble in working with it.
Second, they tried to improve the production of β-carotene with redundancy, adding a second copy of the gene for the enzyme catalyzing the first step of the pathway. Initially they relied on the baker’s yeast’s natural enzyme, BTS1, to carry out the conversion of farnesyl diphosphate to geranylgeranyl diphosphate, but in their work to optimize their system, they found that they could increase the production of β-carotene if they gave its production a kick start by introducing some redundancy to this first step of the pathway. They imported another enzyme from the red-colored yeast, crtE, to help convert farnesyl diphosphate to geranylgeranyl diphosphate. Their final system, then, relied in part on redundancy for reliability. It contained two genes, crtE and BTS1, that both make enzymes that carry out the first biochemical reaction for β-carotene biosynthesis despite their entirely different DNA and protein sequences.
Finally, the researchers improved the system with one additional modification. They determined that they could increase β-carotene production by including a second copy of the crtI gene. In this case, the researchers suspected that the increased reliability with the extra crtI gene was due to increased expression level of the enzyme rather than the other advantages of redundancy we discussed in the earlier section, such as having an extra copy available in case the first copy fails.
Thanks to all the clever genetic engineering of this VitaYeast strain by researchers, we now have a yeast strain that appears bright orange and that the iGEM team at Hopkins in 2011 could work with. Much to the team’s disappointment, though, the strain that had been engineered for reliability is still not orange 100 percent of the time. Restreaking a single colony of this yeast strain onto a petri dish yields orange colonies but also some red, yellow, and white colonies. BioBuilder’s Golden Bread activity investigates this instability and experiments with ways to improve the strain’s performance.
Additional Reading and Resources
§ Ashby, M.F. (1999) Materials Selection in Mechanical Design [ISBN: 0750643579].
§ Petroski, H. (1985) To Engineer Is Human: The Role of Failure in Successful Design [ISBN: 0679734163].
§ Petroski, H. (1994) The Evolution of Useful Things: How Everyday Artifacts—From Forks and Pins to Paper Clips and Zippers—Came to be as They are. [ISBN: 0679740392].
§ Verwaal, R. et al. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous Appl Environ Microbiol. 2007;73(13):4342-50.
§ Website: Golden Rice (http://www.goldenrice.org).
§ Website: World Health Organization, micronutrient deficiencies (http://bit.ly/micronutrient_def).
Golden Bread Lab
This lab introduces students to the eukaryotic experimental organism S. cerevisiae, which differs from E. coli in growth rate and methods genetic manipulation. With the goal of engineering reliability, this experiment attempts to characterize the strain’s instability, using colony PCR to assess its possible genetic sources. The engineering concept of redundancy is emphasized by transforming the yeast with a plasmid containing a synthetic version of one of the genes in the engineered pathway. You can use the Golden Bread lab procedures to teach biotechnology skills such as solid-state microbial culturing and DNA electrophoresis.
Design Choices
The fact that a single strain of yeast, which is built using one genetic program, produces yeast in a variety of colors suggests that the VitaYeast strain is not producing β-carotene as reliably as biological engineers would like. To investigate the source of the unreliability, we will begin by assuming that the different colony colors result from a genetic instability, a change to one or more of the genes in the β-carotene metabolic pathway that occurs as the cells divide. You will conduct experiments to quantify the extent of the unreliability and assess where the genetic change occurred.
In these activities, you will focus specifically on the crtYB gene, because it is the only part of the system that does not have any genetic redundancy already engineered into the pathway for β-carotene production. It is important to recognize, however, that there could be other explanations for the unreliability, including the action of genes not primarily involved in the β-carotene pathway, or completely nongenetic causes, such as differences in the growth medium. Although we will not investigate these other potential sources of unreliability here, they represent excellent follow-up experiments.
Experimental Question
We will investigate, and attempt to fix, genetic instability in the VitaYeast strain using a tried-and-true engineering strategy of redundancy. By focusing our attention on one specific gene, crtYB, we effectively narrow our experimental question to one that is straightforward and testable: will addition of a synthetic crtYB gene, crtYB’, to the VitaYeast strain reduce, or eliminate entirely, growth of non-orange (that is, red, yellow, white) colonies?
To address this question, we will need to compare two values:
§ The number of red, yellow, and white colonies relative to the orange colonies, or the percent of non-orange colonies, before addition of crtYB’.
§ And the percent of non-orange colonies after addition of crtYB’.
If the percent decreases, we can conclude that redundancy did, in fact, reduce genetic instability in the system. If the percent stays the same, perhaps we should try adding a different gene or genes or take another approach entirely. And if the percent increases, we might question whether the procedure we used to introduce the synthetic gene had unanticipated side effects.
By thinking through possible outcomes to the experiment, we can better design the necessary controls. For the transformation procedure, which introduces the new copy of crtYB on a plasmid, we can imagine using both negative and positive controls. A negative control for transformation is a sample that we do not expect to grow on our selection plates, in this case plates that lack an essential amino acid, because it does not contain a plasmid with the selection marker, a gene that makes the same essential amino acid. To prepare this sample, we will need to take it through all the same steps as the experimental sample except no plasmid DNA will be added. If the negative control does grow, contrary to our expectations, we know that something about the procedure or plates is not effectively separating transformed cells from cells without plasmid DNA.
A positive control can help us address questions about unanticipated side effects from the transformation procedure. We expect the positive control to grow on selection plates but do not want it to contain the extra copy of crtYB (that would make it the same as our experimental sample). So, for our positive control, we will transform the yeast with “empty” plasmid that contains the selectable marker but no copy of crtYB. Now, let’s return to our “thought” experiment: what if the genetic instability increases post-transformation? By comparing to the positive control, we can determine whether crtYB’ or the transformation alone is causing this unexpected result. In fact, transformation is a harsh procedure that can cause unintended changes to yeasts’ DNA, so it is important to rule this out as a possible source of genetic instability.
Our experimental sample will contain VitaYeast and a plasmid containing both the selectable marker and the crtYB gene. We will compare the percentage of non-orange colonies to those on the positive control plate post-transformation. This will provide the most direct, apples-to-apples comparison and make it possible for us to attribute any change in color to the crtYB’ gene itself, not any other confounding factors.
Getting Started
You will receive a strain of yeast that already contains the engineered β-carotene pathway. You will begin by conducting your own simple experiment to measure the reliability of β-carotene production by restreaking the orange yeast and counting the numbers of orange, red, yellow, and white colonies.
You will then begin working with one or more of the non-orange colonies. You will conduct a PCR experiment to determine whether the crtYB gene is still present. Recall that crtYB is the only gene in the system that has no redundant copy. We will provide the primers to amplify crtYB, but you may also design primers yourself for this gene or for the others in the pathway.
We will also provide you with a plasmid containing a redundant but synthetic copy of the crtYB gene, which we call crtYB’. This gene produces the same protein product as the original crtYB gene but encodes it with a codon-shuffled DNA sequence. This recoding of the gene can happen thanks to the degeneracy of the genetic code. As with the PCR experiment, you may also go through the exercise of designing this redundant copy yourself. You will transform this redundant copy of crtYB into the red, orange, yellow, or white yeast. The transformation protocol differs slightly from that for transforming E. coli, but the basics are the same. After plating the transformation reaction and allowing the strains to grow for 2 days, you will count the number of colonies of different colors to assess whether introducing this redundancy has made the system perform more reliably.
The yeast strain for this experiment contains the β-carotene-producing genetic circuits integrated into its chromosomal DNA. The yeast will arrive as a “stab” or “slant,” a test tube with a small amount of yeast on a slanted media.
1. Streak out the yeast onto a petri dish using a sterile toothpick or inoculating loop: gather a small amount of cells from the stab on the toothpick or loop, and transfer the cells to a petri dish containing Yeast extract Peptone Dextrose media (YPD) agar.
2. Place these petri dishes media side up in a 30°C incubator for 36-48 hours. Plates can also be kept at room temperature incubation, but cells will grow more slowly.
Lab Protocol
This is an abbreviated version of the lab protocol and includes only the transformation activity. Instructions for colony PCR and designing a redundant synthetic gene are available on the BioBuilder website.
These instructions are using a yeast transformation kit to prepare competent cells. The contents of the kit are proprietary but are most likely the ones you’d find if you googled chemically competent yeast cells.
Transforming VitaYeast
1. For each transformation you want to perform (positive control, negative control, experimental), label a microfuge tube. Then add 500 μl of water and swirl a toothpick full of vitamin A-producing yeast in each tube.
2. Harvest the yeast by spinning the tubes for 30 seconds in a microfuge.
3. Remove the supernatant from each pellet by aspirating or pipeting it away into a waste beaker with some 10% bleach in it. You do not have to remove every drop of the supernatant.
4. Wash each pellet of cells by resuspending them 500 μl of “wash solution” or “solution 1” (most likely just sterile water!) from the kit.
5. Harvest the cells in a microfuge, spinning 30 seconds at full speed.
6. Aspirate or remove the supernatant as before.
7. Resuspend each pellet in 50 μl of “competent solution” or “solution 2” (most likely lithium acetate and DTT, which permeabilizes the yeast through an unknown mechanism). Unlike chemically competent bacteria, competent yeast are not “fragile” in this state and can remain at room temperature.
8. Add 5 μl of water to the negative control tube. Flick the tube to mix the contents. This tube contains no DNA.
9. Add 5 μl of pRS414 DNA (50 ng) to the positive control tube. Flick the tube to mix the contents. This plasmid bears a yeast origin of replication and a TRP1 gene and will serve as a positive control for transformation.
10.Add 5 μl of your pRS414+crtYB’ DNA (50 ng) the experimental sample tube. Flick the tube to mix the contents.
11.To each tube add 500 μl “transformation solution” or “solution 3” to your cells. This material, most likely polyethylene glycol (”PEG” aka antifreeze) is thick and goopy and is included in transformation protocols to help deliver the DNA into the yeast. Use your P1000 to pipet the solution up and down with the yeast, and gently vortex the tube to make an even suspension.
12.Incubate the tubes at 30°C for approximately one hour, along with an equal number of -TRP plates with their lids ajar to evaporate any moisture on their surface. During this hour you can periodically “flick” your tubes to mix the contents; this will help keep the cells from settling to the bottom.
13.After at least an hour, flick the tubes to mix the contents and then spread 250 μl of each mixture on an appropriately labeled -TRP plate.
14.Incubate your petri dishes, media side up, at room temp or in a 30°C incubator for two days.
15.After you return to collect your data, determine the number and color of colonies on each dish.

