The Illustrated Insectopedia - Hugh Raffles (2010)
Vision
1.
Academy Studios, an exhibition design and fabrication company based in Novato, California, created these interactive stations for the Arthropod Zoo at the North Carolina State Museum of Natural Sciences. They built a seven-foot praying mantis and a dragonfly with a twelve-foot wingspan—both anatomically accurate!—but it’s these masks that get the most attention, spooky-looking sci-fi helmets that, as Academy’s promotional materials put it, “give visitors a chance to see life through a bee’s eyes.”

Robert Yagura, then Academy’s creative director, told me that they used hexagonal pieces of Lucite to mimic the facets of a bee’s compound eye and bonded them to a curved form to produce a fractured image. But even with that prosthetic, Robert told me, the visitor does not see like a bee. For a start, a bee’s sensitivity to the electromagnetic spectrum is shifted significantly to wavelengths shorter than those visible to a human. At the low end, below 380 nanometers, bees are able to make out ultraviolet light invisible to us; at the high end, they’re red-blind, red appearing to them as an empty blackness, the absence of light.
The little-remembered zoologist Charles Henry Turner shares credit with Karl von Frisch for providing the first glimpse of the world through the eyes of a bee.1 Turner, the first African American to receive a Ph.D. from the University of Chicago and the author of more than fifty research papers, published his account in 1910, at the beginning of a long career as a science teacher in public high schools. Von Frisch completed his studies in 1913, well before he became director of the Institute of Zoology at Munich and witnessed the honeybees’ dances. He was already driven by the impulse to demonstrate the capacities of his tiny friends that would eventually win him the Nobel Prize. Despite the extravagance of floral color and the intricate economies of codependence that tie insects and angiosperms across the millennia, before Turner and von Frisch had given the matter their attention, it was commonly thought that insects were entirely color-blind.
Von Frisch’s refutation is famous—and characteristically and elegantly low-tech. He set out a series of dishes on cards. In just one square, the only blue card in a field of varying grays, the dish contained sugar water. He began by training his bees to visit this card and dish. Then, over several hours, he varied their position in the matrix. Next, he removed all the cards and dishes, replacing them with a new set of identical materials, only now leaving the dish on the blue card empty. As he expected, the bees returned to the blue card, attracted by color rather than odor or location.2 As von Frisch explained, this behavior demonstrated the bees’ “true color sense,” not simply their ability to distinguish among light intensities. If their vision were monochromatic, he pointed out, they would have found at least some of the gray cards indistinguishable from the blue.3

There’s little argument nowadays that most insects see in some form of color. Through electrophysiologic experiments on photoreceptive cells, researchers can readily demonstrate the capacity for color vision. They know, for example, that bees, like humans, are trichromatic, possessing three types of photosensitive pigments that absorb maximally in different parts of the spectrum (though in green, blue, and ultraviolet rather than our red, green, and blue). And they also know—though there are few ways of conceiving what it might actually mean—that dragonflies and butterflies are often pentachromatic, possessing five types of pigments. (They also know that mantid shrimp have receptors sensitive at twelve different wavelengths!)
It is one thing, however, to demonstrate that animals possess the capacity for color vision, quite another to show that the world through which they move glistens and glimmers, as does ours, in multiple hues. For this, researchers rely on behavioral studies, and they still use the techniques pioneered by Turner and von Frisch, training their animals to respond to food rewards and colored patches.
But insects can be recalcitrant research subjects, and so far this type of work has been carried out only among honeybees, blowflies, and a few species of butterflies.4 Given the distinctive absorption spectra of these animals’ photoreceptors, we can be fairly certain that objects will appear quite different to them than they do to us. Many flowers, for example, look quite different when viewed through an ultraviolet filter. These black-eyed susans (Rudbeckia hirta) display a bull’s-eye pattern that seems to guide bees, wasps, and other pollinators to their target; other flowers have a characteristic runway pattern leading to the same destination.

This is so simple yet so intriguing. There are invisible worlds all around us, parallel worlds. Familiar objects have secret identities, some of which we can access through straightforward mechanical tricks, like Lucite fractals and UV filters, but others of which remain inaccessible, even to our imagination (twelve pigments?). We pass through not just purblind but encumbered by the everyday assumption that the world we see is the world that is. In this respect at least, our perception is rather shallow, though I admit it’s unlikely bees or butterflies are any more decentered.
Nonetheless, at the very least, the natural world’s indifference should make us wary of assuming too quickly that flowers that draw our eye are similarly seductive to a pollinator. Such hidden truths make visible one important fact about vision (our own and that of other beings): it is a property not only of the viewer and the object but also of the relationship between them.5
2.
The closer we look, the more we see. Bee masks and UV photos are not just intriguing; they’re beguiling. If we could only re-create an insect’s visual apparatus, they promise, we could see what it sees, and if we could see what it sees … why, then we could see as it sees, too. But I doubt many of us, including scientists and exhibit designers, believe this. Vision is so much more than mechanics.
The Soviet entomologist Georgii Mazokhin-Porshnyakov drew attention to this long ago: “When we talk about vision,” he wrote in the late 1950s, “we imply not only that animals are able to distinguish objects visually (i.e., the stimuli), but also that they are able to recognize them.”6 Photoreception, on its own, he suggested, is of little value to an organism; what counts is the ability to identify an object and make some sense of it. Reception presupposes perception. Insects see with their brain, not their eyes.
In this respect, an insect’s vision is identical to that of a human. Like ours, an insect’s vision is a complex sorting procedure, a way to filter and hierarchize objects in the world, one sense among several interdependent senses, one entangled element of perception.
Frederick Prete, a biologist at DePaul University who studies the visual universe of praying mantises, points out that until quite recently the standard scientific assumption was that insect vision operated by exclusion, that bees, butterflies, wasps, mantises, and similar creatures were designed “to ignore all but some very limited, specific types of visual information … [such as] a small, moving, fly-shaped spot just a few millimeters away … [or] yellow flowers of a certain size.” Instead, Prete and his colleagues demonstrate, mantises and many other insects deal with sensory information in ways not dissimilar from those of humans: “they use categories to classify moving objects; [and] they learn and use complex algorithms to solve difficult problems.” Prete describes human processing of visual information as a type of taxonomy:
We filter sensory information by recognizing and assessing certain key characteristics of the events and objects around us, and we use that information to identify an event or object as an example of a general class of events or objects. For instance, you would not reject a meal … because it did not look like a specific, idealized plate of food. You would assess its characteristics (odor, color, texture, temperature), and if they all met certain criteria, you would take a bite. In this case, the novel meal is an example of the category “acceptable meal.” Likewise, we can learn that a particular task—mending a ripped curtain, for instance—is an example of the category “sewing material together.” So, when attempting to mend a curtain for the first time, we apply the rules that we learned are successful in other, analogous mending tasks. In other words, we have acquired and employ an algorithm, or “rule of thumb” for solving specific problems of this general type.7
A mantis, write Prete and his colleague Karl Kral, is confronted by a large number of potential meals in the course of a day, and like us, it both creates and deploys a relational category (“a theoretical, perceptual envelope”) that corresponds to the thought “acceptable meal.” To evaluate an object, the animal draws on experience—learned from past events and encounters—to assess a series of “stimulus parameters” that include the object’s size (if it is compact), its length (if it is elongated), the contrast between the object and its background, the object’s location in the mantis’s visual field, the object’s speed, and the object’s overall direction of movement.8 A varying number of these criteria must be met for the mantis to strike. Yet, rather than a reaction being triggered by the meeting of a specific threshold, the mantis takes into account the relationship among different data in each parameter. Kral and Prete call this calculation a “perceptual algorithm” (and make the not-unreasonable point that if it were described in primates, it would be considered abstract reasoning).
Along with a small number of other invertebrate scientists who integrate behavioral and neuroanatomical studies in what is sometimes called psychophysiological research (that is, research on the connections between the psychological and physiological aspects of behavior), Kral and Prete write un-self-consciously of the complexity of insect behavior, of the correspondence between the ways insects and vertebrates (including humans) make sense of the world, and of the insect’s mind.
But maybe these insects are just a little too calculating, modeled a little too much on the rational actors of classical economic theory (who we know from our own experience don’t really exist). Maybe they’re not lively and spontaneous enough. How do we know they always calculate solely according to the logic of the hunter? Might they have other desires? Or maybe this is exactly how mantises are even if we don’t have to assume that butterflies, say, or fruit flies proceed in this way too. No matter; this is thought-provoking work: there is a cognition here, say Kral and Prete, that is dependent on but somehow not reducible to physiology. Yet, if cognitive processes are irreducible to electrochemical function, what exactly are they? Nobody seems very sure.9
It’s worth noting that these questions are central to contemporary neuroscience, the interdisciplinary field concerned with the study of the nervous systems of animals. Neuroscience is dedicated to physiological explanation but is nonetheless deeply preoccupied with questions of mind, with such indeterminate phenomena as consciousness, cognition, and perception, with material solutions to what many might consider ontological or even metaphysical problems. In neuroscience, it is axiomatic that the brain is the center of all animal life—“the key philosophical theme of modern neural science is that all behavior is a reflection of brain function” begins a standard reference work.10 “Higher-order” brain functions, such as metacognition (thinking about thinking) and emotion, tend to be understood as functional outcomes of brain anatomy and physiology.11
Yet the model of perception that overlies this straightforward if controversial principle is formidably elaborate. Perception is conceived as a set of dynamic, interactive brain functions that integrate cognition and experience and include filtering, selection, prioritization, and other forms of active and flexible information management in the context of previously unimagined neuroplasticity. One example might be the interplay between the brain and particular visually salient objects, such that the objects instantly and without conscious registration are isolated within a saturated and nonhierarchical visual field. Such ideas are fully congruent with the type of perceptual algorithm developed for insects by Kral and Prete (and others; see, for example, the two decades of work on honeybee cognition carried out by Mandyam Srinivasan and his team at Australian National University). Yet this parallelism between people and invertebrates would, I suspect, seem foolish to many neuroscientists, for whom the marvelous size and complexity of the modern hominid brain—specifically the number of its neural connections—is the decisive marker of human exceptionalism.
Kral and Prete are likely to find still less support in the social sciences and humanities, though for different reasons. Research on vision here emphasizes the role of culture and history in mediating between the human eye and the world.12 For cultural analysts, physiology often provides little more than a set of possibilities for a complex human perceptual engagement with the world. How humans see and what we see are understood as profoundly shaped by social and cultural history. Vision, and perception more generally, are neither unchanging in time nor constant across cultures.13 They have history—several histories, in fact, as the character of perceptual understanding is understood to be shaped by regional and national aesthetic cultures. Key moments of transformation are tied to the emergence of specific visual technologies. In the West, for example, among other moments, scholars have drawn attention to the invention and dissemination of linear perspective in the fifteenth century and the shift in the nineteenth century to a preoccupation with surface morphologies: the preoccupation with the superficialities of objects and bodies with which we still live.14 In these accounts, vision—the ways in which we observe people and things, the forms of categorization that are embedded in our own ways of looking, and the technologies through which we, in turn, are seen, surveilled, classified, assessed—is central to the ways in which we understand ourselves and are understood by others; it is a source of culture-history-society as well as its outcome.
Such a different vision of vision! Unlike the isolated neurobiological brain, the social brain is immersed in a world that is itself overflowing with meaning, is deeply part of a universe in which even so-called natural phenomena are always simultaneously both biophysical and cultural-historical, so that colors, for example, are always at the same time both measurable wavelength and shimmering story (in which we can’t escape knowing that pink—whether it works for us or not—is cuter than navy blue). In this vision, people learn to see, and the form and content of that learning are specific to time and place. A man blinded years before recovers his sight and must be taught to recognize perspective in culturally effective ways; a woman leaves the closed forest in which she has spent her life and has to make radical, even traumatic adjustments before understanding the spatiality of the urban landscape that is now her home.15
Yet history, politics, and aesthetics—the central categories employed by cultural theorists to explore vision—are by definition exclusively human, are, in fact, definitively, classically, human. Though their proponents may disagree about almost everything else, when it comes to human exceptionalism, there is an emphatic alliance between the social brain, with its immersion in culture, and the neurobiological brain, preoccupied as it is with size and physiological complexity. And the differences on which these competing visions converge are surely not trivial. How can we hold on to them and at the same time refuse their implicit hierarchy?
3.
“The best of the [insect] eyes,” wrote the optical-instrument maker Henry Mallock in 1894, “would give a picture about as good as if executed in rather coarse wool-work and viewed at a distance of a foot.” Indeed, Mallock continued, a compound eye with the resolution of a human eye would itself be a spectacle. Mallock estimated its diameter at more than sixty feet.16 Why so monstrous? Because to adequately counter diffraction—the tendency of light to spread out and blur as it passes through a narrow opening—each lens in every one of the many facets of the compound eye would require a diameter of 0.08 inches, the size of a human pupil—small, perhaps, but an eightyfold increase for a bee.17
Mallock’s fantastic notion—the insect head: outsize, outrageous, but not horrific, not Cronenberg’s fly—makes me want to climb back into one of those Lucite masks! Even though I know that they don’t really work, that there’s far more to vision than this, the urge to see through another’s eyes isn’t easily suppressed. And I’m far from alone. So many people have been driven to try it, the more scientifically minded concocting ingenious ways to record the view directly, delicately scraping out the eye’s internal structures, removing the retina, cleaning the cornea, experimenting with light, microscopes, cameras, the product less immersive than a mask but more objective seeming, more authentic feeling. This impulse to capture the vision of another being is potent, and I believe I’m right in thinking it derives its power from the unusual coincidence it creates between the two visions of vision we’re caught in here: the promise of the natural sciences (that is, the revelation of how things work, a revelation of structure and function that is often ultimately rather unrevealing) and the inaccessible dream of the human sciences (the utopian dissolution of ontological difference, the impossible yearning to enter another self). The most recondite mysteries are resolvable, the impulse tells us. Everything can be illuminated.
Anton van Leeuwenhoek, the discoverer of bacteria, sperm, and blood cells, of the mouthparts and stings of bees, of animal motion in a droplet of water, and of many, many other microbiological phenomena, was the first to see the gleam in a compound eye. Shining a candle through an insect’s cornea, he used one of the gold and silver compound microscopes of his own invention, one of the microscopes sold by his family after his death and now lost, one of the microscopes that Robert Hooke copied to access the unimagined and deeply disturbing world he laid bare with his draftsman’s precision in Micrographia, the volume that includes his famous engraving of a dragonfly’s head—its diabolical masklike face made visible for the very first time—and in which he recorded his disbelieving observation that reflected perfectly from each of the facets of the animal’s compound eye was “a Landscape of those things which lay before my window, one thing of which was a large Tree, whose trunk and top I could plainly discover, as I could also the parts of my window, and my hand and fingers, if I held it between the Window and the Object.”18
Hooke wondered aloud about the optics of his “drone-fly” (“How exceeding curious and subtile must the component parts of the medium that conveys light be, when we find the instrument made for its reception or refraction to be so exceedingly small?”),19 but it was van Leeuwenhoek—thirty years later—who first realized that the image transmitted to the fly’s brain was fractured, that each facet of the eye captured its own image. Van Leeuwenhoek recorded his account in a breathless letter to the Royal Society of London, published in 1695, an age when the arts and sciences were still negotiating their formal separation. “What I observed by looking into the microscope,” he informed his colleagues, “were the inverted images of the burning flame: not one image, but some hundred images. As small as they were, I could see them all moving.”20
Almost exactly two centuries later, Sigmund Exner, the eminent physiologist who would guide his young nephew Karl von Frisch in setting up the family natural history museum on the shores of Lake Wolfgang, was completing The Physiology of the Compound Eyes of Insects and Crustaceans, the first authoritative account of insect vision and a groundbreaking monograph whose claims survive intact to this day.21 Exner was an assistant to Ernst Brücke, the professor of physiology at the Institute of Physiology in Vienna who persuaded Sigmund Freud to reject neuroscience for neurology. Exner and Freud were colleagues at the institute, both under the mentorship of Brücke, and like Freud, Exner was at the same time subject to the compulsion of vision and fascinated by its mechanics. With great care and labor, he succeeded in taking a photograph through the compound eye of the firefly Lampyris, but the image he saw was quite different from van Leeuwenhoek’s.
How could the multiply-fractured compound eye, with its numerous facets, produce only one image, and how could that image be erect rather than inverted, like the image that travels to the brain from the eye of both the fly and the human being?

Although the difference is not externally evident, Exner knew that there are two distinct types of compound eyes. The compound eye of the fly examined by van Leeuwenhoek is made up of multiple light-gathering units, called ommatidia, each of which is itself a single, independent eye that collects light from just a narrow section of the animal’s field of view. Exner discovered that in this type of compound eye—called an apposition eye—light passes through the hexagonal facet lens into the crystalline cone, with its sheath of pigmented cells that block ambient light from adjacent ommatidia, down the cylindrical light-sensitive rhabdom, which houses the eight photoreceptive retinal cells, and all the way to the nerve cells that transmit the image to the optic ganglion and into the brain, where the inverted mosaic produced by the retinal cells is turned into a single erect image.
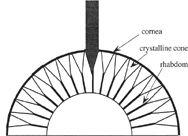
But Exner also knew that like moths and many other insects that fly at twilight and after dark, the firefly, whose retinal image he reproduced in his 1891 monograph, is nocturnal and possesses what is called a superposition eye, an instrument 100 times more light sensitive than the apposition eye of diurnal insects.
Rather than being separated into individual ommatidia, the retina of the superposition eye is a single sheet, and it lies deep in the eye, above a clear zone across which light is focused. We might say that in the superposition eye the ommatidia cooperate: the image falling on its retina at any point is the product of multiple lenses.22
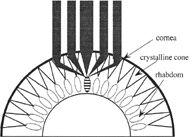
But the real puzzle was how such optics succeeded in producing an erect image. And it was Exner, working on this problem throughout the 1880s without tools for definitive demonstration, who figured out that the rhabdom of the superposition eye functions as a two-lens telescope, redirecting the rays of light so they cross each other within the cylinder and reverse the image. “Clearly,” says the biologist Michael Land, “we are dealing here with something quite out of the ordinary.”23 The views below, taken by Land and Dan-Eric Nilsson, demonstrate the difference between the images produced by the two types of compound eyes. The inverted apposition view, on the left, was photographed through the cornea of a robber fly; the rather blurry image (of Charles Darwin) on the right is seen through that of a firefly.24

The number of ommatidia in a compound eye varies enormously, from single digits in some ants to more than 30,000 in some dragonflies. As we might expect, the more ommatidia that are present, the better the eye’s resolution. But even the best compound eyes cannot focus, and cannot move in their sockets (and so demand that the insect swivel its entire head to shift its view), and, except at very close range, compound eyes have relatively very poor acuity. Where insect eyes do excel, as anyone who’s tried to snatch a fly or squish a mosquito knows very well, is in their sensitivity to movement. Flying insects in particular often have an extraordinarily wide field of vision, as great as 360 degrees in those dragonflies whose eyes meet at the top of the head.
But this is not the insects’ only basis for motion detection. To compensate for the animals’ accelerated flicker-fusion frequency—the rate at which a moving image appears as one continuous flow rather than a series of discrete events like those on the pages of a flip-book—a movie designed for (or by) a fly would run five times faster than the standard twenty-four frames per second. Flies, this suggests, live in a world that moves much faster than ours. They are born and they die within days, weeks, or months, not decades. They occupy a different plane from us. Not only is it a plane in which acuity, pattern, and color are distinctive, but it is also one in which space-time is lived in another register. If we think of our senses as mediating our relationship with the world around us, we might wonder about the perceptual, intellectual, and emotional lives of beings, including human beings, whose sense organs differ from our own. Some of this mystery can be captured by blurry pictures and plastic masks. Some of it should be allowed to throw our own perceptual certainties into relief.
4.
This thought leads to one more view from an insect’s eye. And this time it’s not a photograph. It’s a different kind of re-creation, and it was made in the 1930s by the great Estonian biologist-philosopher Jakob von Uexküll. In the forests through which von Uexküll wanders, all sensate beings are subjects, occupying their own Umwelt, an environment defined by the limits and possibilities of their senses.25 All beings live in their own time worlds and space worlds, too, distinct worlds in which both time and space are subjectively experienced through sense organs that differ radically among beings and produce radically different experiences. “The subject sways the time of his own world,” von Uexküll writes. “There is no space independent of subjects.”26
Above is a room as sensed by a housefly. Von Uexküll divides it into “functional tones.” Everything except the plates, the glasses, and the lamp is a “running tone,” a surface on which the fly can run. The heat of the light pulls the animal in; the food and drink on the table hold fast its taste-bud-equipped feet. I don’t believe that the universe of a fly is quite this bland, but nonetheless there is an important insight here. Remember the black-eyed susans? “There can be no doubt,” von Uexküll writes, “that a fundamental contrast prevails everywhere between the environment which we see spread around animals, and the Umwelten that are built up by the animals themselves and filled up with the objects of their own perception.”27
Parts of these alien Umwelten arise from simple motor responses, the kind of thing that Jean-Henri Fabre called instinct. But others are the result of trial and error, of judgment, of “repeated personal experiences.” These are “free subjective products,” and like time and space, they are experiential and individualized.28 This concept doesn’t seem too hard to grasp: that the world is multiple and is different for different beings; that our world is our world and theirs is theirs; that when we meet, it is across and between distinct, intersecting realities. Actually, isn’t this the road we began to travel when we donned the Lucite mask? And now that we’re here, doesn’t it look as if these masks are finally less about the promise of communication and communion, about seeing as the animal sees, than they are about the fact of unbridgeable difference?
Well, continues von Uexküll, hammering his point home, although things exist objectively in the physical world, they never appear in any being’s Umwelt as their objective selves. All animals, ourselves included, know these objective things only as perceptual clues with functional tones—and “this alone makes them into real objects, although no element of the functional tone is actually present in the original stimuli.” And so he continues, on and on, getting deeper and deeper into the flow of his argument (so deep, in fact, that it is hard not to keep quoting him), “we ultimately reach the conclusion that each subject lives in a world composed of subjective realities alone, and that even the Umwelten themselves represent only subjective realities.”29 All of us, all the people and all the animals, live in worlds of our own making, more or less complex, more or less stimulating, similarly subjective.
And then, as if he hasn’t traveled quite far enough, von Uexküll takes an unexpected turn. The worlds of animals and men, he says, are often governed not by logic but by magic. The intricate boring of a bark beetle beneath the surface of a tree is a magical phenomenon. The owner of a dog is a magical figure to the animal. The unlearned routes of migratory birds are similarly beyond understanding. He shows us how an oak tree is many different things to the many different animals that live in and around it. He shows us how sound waves are a different entity to the physicist who studies radio frequency than they are to the musician. (“In the one there are only waves, in the other only sounds. Yet both are equally real.”)30 I think of Annmarie Mol’s atherosclerosis, that gesture of covering the corpse’s face; I think of those strange fighting fruit flies with their poor ground-up heads. “So it goes on,” says von Uexküll. And so we follow him into a universe saturated with signs, a semiotic universe of subjective responses and near-limitless human and animal subjectivities.
Of course I like this. But it makes me nervous. Like a plunge into the void. There is so much possible between seeing and perceiving. And a world of signs is also a world of communication. Senses combining, working together, overlapping, contradicting. So what’s this I’m hearing? Magical sounds from unearthly sources? Sounds? Noise? Music? It’s very loud. And it’s very strange. It’s on my headphones. It’s coming from New Mexico …