How Plants Work (2015)
7 Night Shifts and Other Unexpected Movements
SOME OF MY FAVORITE spring bloomers are tulips—all colors and forms, from the prim species tulips to the drama queen hybrids. My aesthetic side loves the simple yet bold stalks and leaves topped by the perfectly symmetrical flower, and my scientific side loves the fact that every morning they’re closed up tight, opening with the sun and closing again at night. This simple garden flower demonstrates, day after day, that plants move.
We’re used to thinking about our garden plants as rooted in place, and of course they are. But they can burrow, crawl, twine, capture, and strangle as well. Some plants are lightning quick, whereas others need time-lapse photography to show they’ve moved at all. These movements may result in permanent changes to a plants appearance or they may be fully reversible. Let’s find out how and why plants move.

Tulips open and close in response to sunrise and sunset.
Nasty Plants
My garden tulips and many other plants close up their floral parts and/or leaves at night. This phenomenon is called nyctinasty (from the Greek words for night and pressing close). So the onset of nightfall causes the petals of tulip flowers to press close together. This protects vulnerable tissues from nighttime snackers, and it may help insulate spring bloomers from freezing damage.
How do plants know when it’s time to close up shop for the night? We’ve already explored how phytochrome allows plants to measure time, and this is another example of phytochrome at work. When the sun sets, red light disappears and phytochrome changes into its alternate form, which triggers the response. Sunrise reverses the process, when phytochrome absorbs red light and transforms into its daytime form.
Nyctinasty is easy to observe in the summer, but the cold short days of winter can show us yet another interesting phenomenon. Now, I’m in a long-term relationship with rhododendrons. I grew up in Washington State, where it’s the state flower, and I studied their cold hardiness for my doctoral research. I’ve had rhododendrons at every house I’ve lived—even in Buffalo! And there’s one thing they do that I don’t see in other garden plants: they roll up in response to cold temperatures.
Rhododendron leaves are thermonastic. When the temperatures are below freezing, they droop and curl longitudinally, so it looks a little like long green cigars hanging from the branches. Many gardeners think this is a wilting response, but the leaves are full of water during this reversible movement. It’s thought that drooping and curling allows the leaves to avoid damage caused by low temperatures, perhaps by creating a warmer cylinder of air inside the furled leaves. Once temperatures rise above freezing, the leaves perk right back up, none the worse for wear.
Gardeners are familiar with both nyctinasty and thermonasty, but even non-gardening types are fascinated with this next nastic movement. People literally cannot keep their hands off sensitive plants (Mimosa species), whose delicate leaflets fold up on themselves when brushed by a finger. This is thigmonasty, or plant closure caused by touch. The advantage to this is obvious: hungry insects alighting on the leaves are quickly dislodged when their landing strips suddenly disappear.

Rhododendron leaves roll longitudinally when temperatures dip below freezing.
But the titan of thigmonasty has to be the Venus flytrap (Dionaea muscipula). I remember my mother buying one of these when I was about seven years old. It sat on our kitchen windowsill, where I would alternately force-feed it dead insects or play with the leaves. Not surprisingly, it didn’t last long, as it probably spent more energy reacting to my pestering than it gained in consuming food.
Plants have other nastic movements, in response to certain chemicals and water, for instance. The relatively rapid response causes many people to think that plants have nervous systems like animals, but of course they don’t. Instead, the mechanism behind flower and leaf closure relies on regulating water movement.
Water Works
Let’s look at a sensitive plant leaf. It’s structured like a feather, and each little green blade is a leaflet. You’ll note there’s a swollen bump at the base of each leaflet and at the base of the entire leaf. This bump is a pulvinus, derived from the Latin word for pillow. Rather than being filled with feathers, however, the pulvini are plumped up with water.
When a mimosa leaf is brushed, a series of chemical changes is triggered inside each nearby pulvinus, and the result of this chemical cascade is that water rushes into some parts of the pulvini and out of others. The asymmetric turgor causes opposite leaflets to fold together. If there’s a great enough disturbance, the pulvinus at the base of the leaf itself expands, causing the entire leaf to fold downward. Essentially, the plant has hidden its tender green parts from insect or other herbivore damage. Once the threat is past, water seeps from the pulvini back into the surrounding tissues, the leaf petiole rises, and the leaflets unfold.
Venus fly traps also close as a result of insect invasion, but rather than avoiding the bugs, fly traps love to get to know their visitors. The blood-red inner surface of each of the trap leaves has three trigger hairs. Curious insects crawling around will eventually trip one of these triggers, but the trap isn’t sprung until two of the triggers are brushed within seconds of one another. Immediately the trap closes, the soft spiny projections on the edges of the leaves interlocking with one another like clasping fingers. The insect is held in this photosynthetic prison, which continues to close until the leaf edges, and the insect’s doom, are both sealed.

The pulvini of a mimosa leaf are under hydraulic control.
Why does it take two triggering events for the leaves to close? We need to consider the energy that active trap plants expend in closing and opening their leaves. It’s always possible that something inedible could trip a trigger hair—some bit of debris blowing in, for instance—and the plant would waste resources closing on a nonexistent meal. So Venus fly traps have evolved this failsafe mechanism to ensure that trap closure will deliver the goods.
Though not as dramatic, the same changes in turgor pressure that capture hapless insects also cause tulip flowers to close at night. Likewise, our chilly rhododendron leaves droop as both temperature and turgor pressure drop. All nastic movements are rapid, reversible, and nondirectional. In other words, the open-and-close response is always the same regardless of where the stimulation occurred. For instance, you can stroke a sensitive plant leaflet on its upper or lower surface, from tip to base, and the closing response is identical.

Venus fly trap leaves will only close when two trigger hairs are disturbed in quick succession.
Chasing the Sun
Tropisms are another group of plant movements that react differently to environmental stimuli. Tropic (pronounced with a long o) is derived from the Greek word for turn, so tropic movements cause a plant to turn toward (a positive tropism) or away from (a negative tropism) some environmental factor. Some of these tropic movements are quick and reversible like nastic movements, whereas others are slow and irreversible because they trigger a change in the plant’s growth.
What gardener doesn’t love something about sunflowers? Okay, maybe they’re too big and rangy for your yard, or perhaps the yellow-and-black color scheme isn’t to your taste, but they’re easy to grow, produce prodigious amounts of edible seed, and the young flower heads and leaves track the sun. As sunflowers mature, this sun-worshipping behavior stops as the flower stalks stop elongating and toughen up. They’re now stuck in place.
Solar tracking has the lovely formal name of heliotropism. Many garden plants have leaves that follow the sun across the sky. At night, the leaves rotate 180 degrees to face the sun rising in the east. Not many garden flowers actually track the sun, unless you happen to live in an alpine or arctic region. It’s in these cold regions where floral heliotropism is most common. By following the sun in these cold climates, the cuplike flowers collect heat, drawing in pollinating insects for a quick warm-up before they depart with their cargo.
In contrast to these sun lovers, plants in decidedly hotter environments often avoid the sun through the very same process. Rather than tilting their flower and leaf surfaces toward the sun, sun avoiders remain parallel to incoming sunlight. These plants are paraheliotropic and they actively reduce the heat load and drought stress that too much absorbed sunlight can bring.


Heliotropic plants like sunflowers follow the sun, whereas paraheliotropic species avoid it by keeping their leaves parallel to incoming sunlight.
Even though solar tracking depends on sunlight, it’s not controlled by phytochrome like nyctinasty is. Instead, solar tracking is controlled by a pigment called cryptochrome (meaning hidden pigment), which absorbs blue light from the sun and uses it as a means to tell time. This pigment sends a signal to the base of the leaf or flower, where a pulvinus works its magic to move the organ to just the right angle. As the sun moves, so do the leaves and flowers, though they stop if clouds block the sun. Once the clouds have passed, movement resumes and actually accelerates until the perfect angle is reached again.
Chasing the Sun, with a Twist
Heliotropism is a fascinating plant phenomenon, but it’s not the only trick plants have up their little green sleeves for capturing sunlight. All gardeners know that sun-loving plants hate being stuck in the shade. If they don’t languish and die, they try to outgrow their conditions. Many an unwitting homeowner has tried to hog-tie a leaning tree, mistaking the unequal growth pattern for root instability. There’s no way to win this wrestling match with nature.
This is phototropism, a directional plant movement toward (or rarely away from) light made permanent by uneven plant growth. Like heliotropism, it’s activated by blue light, but the responsible pigment isn’t phytochrome or cryptochrome but yet another type of pigment called a phototropin found only in the growing tips of plants. When sun-loving plants desperately seek sunlight in a shady world, that’s phototropism at work. You’ll also see it in neglected container plants that have fallen over and been left horizontal for too long.
How do plants grow unevenly? Auxins are responsible for cell elongation. If auxins accumulate more on one side of a growing shoot than the other, the plant will begin to curve to one side. In phototropic plants, auxins accumulate on the shadiest side of the growing tips, increasing the growth of the cells on that side, and creating a bend in the shoot. Plant producers take advantage of this directional response, rotating their plants to create intricate spirals (lucky bamboo springs to mind here). Of course, any new growth will resume a normal pattern once the plant is sold by the production nursery.
Phototropic responses are strong in sun-loving plants, but much weaker in shade tolerant plants and most conifers. That’s why some trees and shrubs lean away from their neighbors, but others don’t seem to mind close quarters. Watch the development of shade in your garden through the years, and selectively prune sun hogs when necessary.
Going to the Dark Side
Most phototropic plants respond positively to light—they grow toward it—but some species grow away from light. As you might expect, roots are negatively phototropic, but why would some shoots grow toward darkness? This fascinating phenomenon is called skototropism (from the Greek word for darkness) and was documented in the tropical forest vine Monstera gigantea, whose name should tell you that the leaves on these suckers are huge! In fact, gardeners will recognize Monstera species as common greenhouse plants. Needless to say, vines with huge leaves need support, and trees are nature’s trellises. Trees also absorb most of the incoming sunlight in the forest canopy, so the darkest areas in a forest will be where tree trunks are stationed.

Plants usually grow toward the light, like this phalaenopsis on a windowsill.
Now imagine you are a Monstera seedling, with a limited amount of resources for survival. You must reach the sunlight before the seed supply of carbohydrates runs out. You can’t see where the nearest tree is, but you can sense the presence of darkness. Growing toward this black hole is the best shot you have for finding a tree. Once a suitable support has been found, skototropic vines become phototropic and reach for the sky like any normal plant.
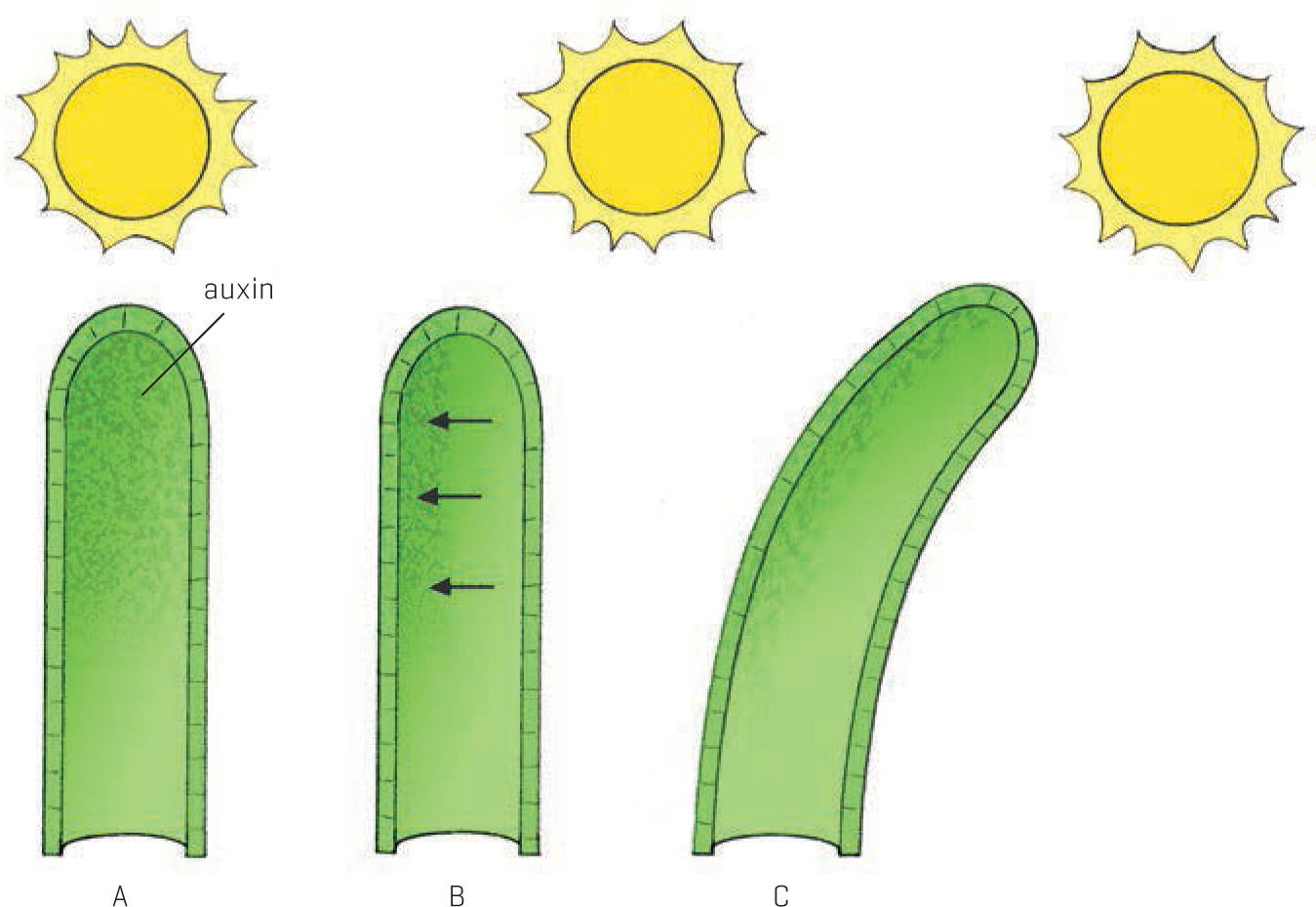
Directional growth is caused by unequal concentrations of auxin in stems. (A) Auxin is distributed evenly in the shoot tip when the sun is overhead. (B) Auxin molecules shift to the shaded side of the shoot when the sun is at an angle. (C) More auxin causes greater cell elongation on the shaded side, so the shoot bends toward the sun.
Skototropism is probably more common than we may realize. Virginia creeper (Parthenocissus species) was recently discovered to be skototropic, and there are doubtlessly other vines that temporarily turn to the dark side. The secret behind how vines twine around their support will be revealed later in this chapter.
Staying Grounded
I mentioned earlier that roots are negatively phototropic, but they aren’t just repelled by light: they’re also attracted to gravitational pull. Gravitropism describes a seedling root’s propensity for growing down (positive gravitropism) and the shoot’s opposite response (negative gravitropism).
The gravitropic response in roots is variable and depends a lot on the plant’s life stage. Seedling roots are strongly gravitropic; they determinedly grow straight down so they can stabilize the seedling and start harvesting water and nutrients. These are taproots, and most plants start life with a deep, strong taproot. But as the seedling develops, it’s more important to start colonizing the surrounding soil. New roots develop that are less and less gravitropic and grow horizontally, attracted more to pockets of water and nutrients than to a gravitational pull.
The means by which roots grow downward and shoots grow upward is the same as with phototropism: unequal distribution of auxin along the growing tips of both parts of the plant. For roots, you’ll find more auxin in the upper side of the growing tip, and in shoots you’ll find more in the lower side. The increased elongation in the first case pushes the growing tip down, whereas in the second it pushes the growing tip up.
But what’s really sensing the gravitational field? It’s not a pigment, because those sense light. In gravitropism, it’s currently thought that tiny grains of starch, called amyloplasts, are the low-tech way that plants figure out which way is down. (It’s similar to the way your inner ear senses gravity, though you don’t have starch grains in there.) The granules are denser than the cytoplasm, so they sink to the lowest point of the cell. Auxin is allocated to the cells in this same side of shoots, but to the opposite side in roots.
Defying Gravity
By and large, the aboveground parts of plants are negatively gravitropic, as their dependence on sunlight for survival overrules almost any other environmental signal. This dogged tenacity is how lucky bamboo (Dracaena braunii) is grown into such interesting shapes. Pots of Dracaena are laid horizontally and regularly rotated, creating a spiral or some other design as the plants continue to grow upward.
Gravitropism is also important in horizontal branch stability. As tree branches increase in girth, they get heavier and are more likely to break. Trees form what’s called reaction wood, and more of this is formed on the bottom half of the branch than on the top. If you were to look at a cross section of one of these branches, you’d see very narrow growth rings at the top of the branch, progressively becoming more massive as you move to the bottom of the branch. Unequal auxin distribution causes the cambium to expand faster on the lower side, creating this elongated bulls-eye effect.
Plant Associations
When you place a plant in the ground, it changes its surrounding environment. Some of the better things plants might do are provide structural support for climbing plants, create shade for sun-intolerant species, increase soil nutrition through nitrogen-fixing bacteria in their roots, or take up and neutralize salts and toxins in the soil. They don’t do it to be nice to their neighbors, they do it as a matter of survival. Unrelated plants in the same vicinity can benefit from these environmental modifications and in turn may provide their own benefits. When both plants benefit, you have what ecologists call a plant association.
Intercropping or polyculture is a gardening strategy that takes advantage of mutually beneficial plant relationships. Together, plant associations increase local biodiversity by attracting and retaining beneficial insects while confusing pests that prefer a monocultural diet. Give a mixed vegetable garden a try! Combine different vegetables with one another to see what works best for your local conditions.
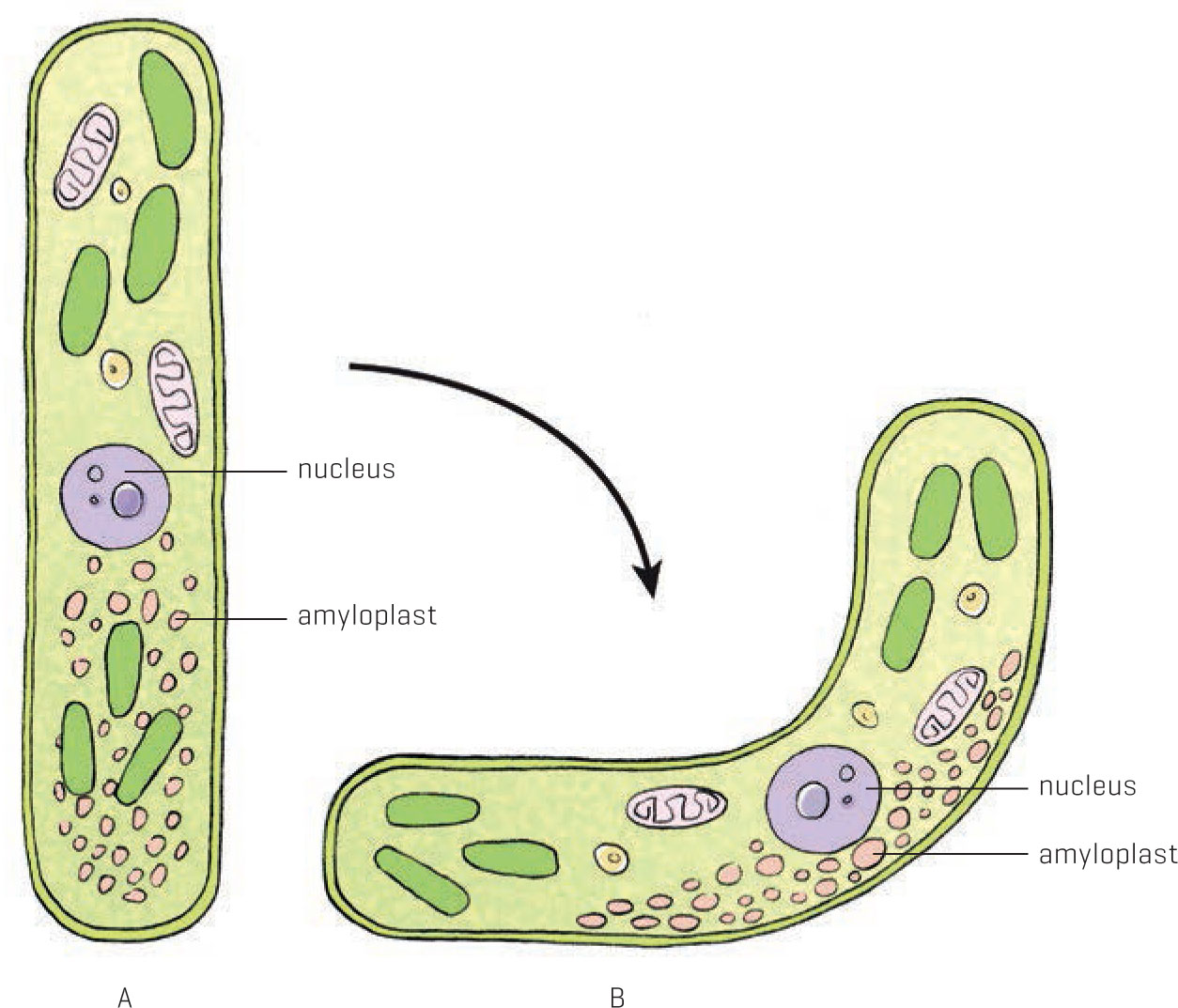
Amyloplasts collected at the base of an upright cell (A) are redistributed when this cell becomes horizontal. The redistribution allows the plant to reorient itself upward (B).
Probably the most bizarre gravitropic behavior is found in peanuts. Once the peanut flower is pollinated, it becomes positively gravitropic: it heads for the ground and buries itself to finish development. This is where the mature nuts are found, thus explaining the alternative name of groundnut.
You might wonder whether positive phototropism and negative gravitropism aren’t just different names for the same thing. They really aren’t. They are sensed by different parts of the plant as a way of fine-tuning its response to light. For instance, when a sun-loving plant is trying to outgrow shade, it will lean toward light (photo-tropism), but not at a 90-degree angle. Instead, it continues to grow upward (negative gravitropism) as well as sideways. If you remember your geometry, the hypotenuse of a right triangle is shorter than the sum of the two sides. So the plant is conserving precious resources by taking the shortest distance between two points, rather than stair-step in its efforts to reach the sun.

Spiraling in lucky bamboo is caused by continually rotating pots under a light source.
That’s all well and good for woody plants with their built-in structural supports. But what about those vines we left back in the heart of darkness?
Tell Me Why the Ivy Twines
In our backyard we have a wisteria vine that snakes itself around a sturdy trellis. Dotted along the fence line are various clematis cultivars, some of which have lassoed branches of nearby trees and shrubs with their tenacious tendrils. Though the floral show is stunning, the nuisance factor is high, too. I’m perpetually pruning unruly vines out of young trees and shrubs that would otherwise be completely overrun. It’s no wonder that some vines, like English ivy and traveler’s joy (an oxymoron if there ever was one), have ended up on noxious weed lists.
The winding of vines and tendrils around other plants and upright structures is called thigmotropism. Like thigmonasty in the sensitive plant, physical contact is the stimulus, which in this case causes a directional growth response. Vines have little ability to support upright growth and use thigmotropism to scramble over their neighbors in search of the sun.
My young ginkgo tree is a poster child for clematis catastrophe during the summer. Whenever a young clematis vine would bump into a ginkgo branch, a tendril seemed to emerge overnight and begin to coil. If we were to measure the auxin levels in the tendril, we’d find the levels to be lowest where the tendril touches the ginkgo branch. The high levels of auxin on the opposite side of the tendril cause the cells to grow longer, so the tendril curves around the ginkgo. This unequal growth continues for several rotations until there’s a nice little spiral. With the vine firmly anchored, new tendrils will seek and capture ever higher branches.
The curious gardener might wonder whether vines grow clockwise, counterclockwise, or both. The internet is full of misinformation on the topic, most of it following the popular Coriolis effect mythology, which dictates that vines in the northern hemisphere grow clockwise and those in the southern hemisphere grow counterclockwise. The reality is that more than 90 percent of viny species twine counterclockwise, forming a right-handed spiral. A minority grows clockwise, and a small percentage follows both crowds. Scientists aren’t sure what the advantage might be for counterclockwise growth, but observations suggest that the right-handed spiral might be tighter and therefore clingier.

Dense conifers make a natural backdrop for flowering vines.

Vines are generally counterclockwise twiners (LEFT), but some are contrarians (RIGHT).
Regardless of which direction they spiral, vigorous vines require serious pruning. So do trees and shrubs, especially those that seem to have their own ideas on how to contribute to your landscape design. What causes plants to grow the way they do?