How Plants Work (2015)
5 Why Leaves Can Turn Red Anytime, Anyplace
I’M A LUCKY GARDENER, because I live in a region with distinct seasons. I actually enjoy storing summer clothes and rediscovering what’s in my winter wardrobe. Likewise, my garden packs away its languid summer greens, replacing them with bold reds, oranges, and yellows. If you’re the fanciful type, you may like the notion of Jack Frost painting the autumn leaves. And indeed, cooler night temperatures speed up autumn color transformation. But wait a minute! Some plants have leaves that turn red in midsummer, some whose leaves are red when they’re young and then turn green, and some whose leaves are always red. Why do leaves turn red, anyway?
Anthocyanins: Yet Another Fine Plant Product
It seems as though plants possess more pigments than an artist’s palette! That’s because light is one of the most important and reliable environmental signals plants receive. Pigments select and collect particular wavelengths of light for photosynthesis and other botanical business. The pigments that reflect and transmit most of the red, blue, and purple colors we see in leaves, flowers, and fruits are anthocyanins, a name that literally means flower pigment. Unlike chlorophyll and carotenoids, nested cozily in the chloroplasts, anthocyanins are water-soluble mobile pigments that can scoot easily within and between cells and tissues.
Anthocyanins are multipurpose molecules. Their mobility in water means they can be used as cellular transport systems. Sugars, metals, and other substances can be hooked on to anthocyanins and zipped off to other parts of the plant that need them. Anthocyanins are also a natural sunscreen: under full sun conditions they can protect chloroplasts from solar overload by siphoning off some of the incoming energy. They’re also powerful antioxidants, meaning they can protect plant cells from all kinds of environmental stressors. This is why nutritionists recommend that we eat so many red and blue fruits and vegetables: these antioxidants are beneficial to us, too.

The green-leaved rootstock of this laceleaf maple could take over the red-leaved scion if new green sprouts aren’t pruned.
Why Aren’t All Leaves Red?
With the advantages that anthocyanins offer plants, the curious gardener might wonder why all leaves aren’t red. To answer this, let’s look at some tree cultivars that have been deliberately selected for their permanent red foliage. There are several flowering crabapples with various hues of red foliage available at most nurseries, and many of these have been grafted onto a green-leaved rootstock. If we were to study these trees in the landscape, we’d come across several with green-leaved suckers threatening to overtake the red-leaved scion. Clearly, the rootstock of these crabapples is more vigorous.
It turns out that anthocyanins are expensive to manufacture. Their synthesis takes chemical energy that the plant could otherwise funnel into growth. Even worse, these pigments can interfere with photosynthesis, further reducing the plant’s energy reserves.
Okay, now it sounds like I’ve just contradicted myself. How can anthocyanins interfere with photosynthesis if they’re not in the chloroplast? Let’s review what pigments do with light. Chlorophyll looks green because it absorbs red and blue wavelengths, reflecting and transmitting the green wavelengths to our eyes. The absorbed light is what’s used for photosynthesis. Red anthocyanins reflect and transmit red light, while they absorb blue and green light. The absorbed blue light doesn’t do any work in the anthocyanin molecule—it’s not making sugars or something else—but it’s not available to the chloroplasts, either. So red leaves aren’t able to use all of the incoming blue light for photosynthesis, and the leaves’ ability to make sugars is stifled. Green leaves naturally have the advantage over red leaves in cranking out carbohydrates. In fact, research on red- and green-leaved Coleus has shown exactly that. And red-leaved cultivars grown in the shade tend to be more green than red.
The Young and the Stressless
Tender, emerging leaves are at the mercy of an often hostile world. Insects wait to feast on them. Sunlight can parboil them. Diseases riddle them until they resemble lacework. One of the ways some plants protect their new leaves is with a healthy dose of anthocyanins. This temporary condition, called juvenile reddening, is usually most intense in leaves exposed to full sunlight. Part of this is because of that natural sunscreen function I mentioned. But there may be another benefit.
Leaves are pretty darn tough when they’re mature. But when they are young and expanding, they don’t have the tough waxy cuticles, rigid cell walls, and the other natural defenses that older leaves do. Not only does this make the leaves easier to attack from the outside, but it also means that water loss is greater, too. We can see this happening in the garden on a hot summer day. The newest stems and leaves on our plants wilt first, because they can’t control the evaporation through their expanding surfaces. If water uptake from the roots doesn’t keep up with demand, then these young leaves lose so much water that their tissues are killed. Usually this happens first at the tips and along the margins of the leaves, where you’ll eventually see browning (or necrosis) as an indicator.
Watering and Leaf Wilt
THE PRACTICE
Interpreting leaf wilt as a signal to add water.
THE SUPPOSED BENEFITS
Leaf wilt means that water levels are low in the leaf, so that must mean they are low in the soil, too.
HOW PLANTS RESPOND
While it’s true that leaf wilt is generally due to lack of water in the leaf, the underlying cause is not necessarily lack of soil water. What leaf wilt does tell you is that the roots are not doing their job in taking up sufficient water. Soil that’s too wet, too dry, too hot, too cold, compacted, or salty can injure or kill roots. Pathogens, insects, and rodents can do the same. Poor root systems—those that circle, kink, and generally tie themselves in knots—don’t take water up well. You, the gardener, realize there’s a problem when the leaves droop like steamed spinach. Root around the roots a little and try to diagnose the trouble. Don’t just turn on the hose!

Leaf wilt can be caused by many factors, not just lack of water.

Browning on the tips and edges of leaves is an indicator of drought stress.
Now consider some common garden plants that show juvenile reddening, such as Photinia and English ivy (Hedera helix). The young, expanding leaves rarely wilt, and I’ve never seen leaf browning from water loss in either plant. In this case, what anthocyanins might be doing is holding water inside the leaves by adhesion. Remember, the general rule is “Water always moves to where water isn’t.” Because red leaves have anthocyanins swimming around in the cellular water, that means they have relatively less water than green leaves. It’s harder for water to evaporate away from red tissues than from green ones.

If water is limiting, new leaves won’t expand to their full size.
A final problem young leaves face is successfully reaching full size. During this brief window of time, the bonds between cell wall fibers are loosened, allowing them to slip past one another. When leaves are expanding, they rely on the power of water—turgor pressure—to force cell wall expansion and reach mature size. If young leaves are not turgid enough, whether it’s from too little water uptake or too much water loss, they won’t reach maximum size. In fact, the presence of notably smaller leaves on evergreen trees and shrubs is a good indicator of drought stress.
Not only can young red leaves retain their water in cellular lockdown, they also attract more water from adjacent tissues. Increasing the water content plumps up young leaves, creating the turgor pressure needed for cell walls to slide past one another and guaranteeing maximum leaf size. Once the leaves have stopped expanding, the anthocyanins disappear and red fades to green.
How Dry I Am, How Red I’ll Be
While some plants have red leaves only during their teenage years, others wear these flashy pigments all their lives. Many of these are species found in environments that are bone dry much of the year.
We’ve already seen that anthocyanins help young leaves hang onto their water during expansion. In the same way, red-leaved trees and shrubs can tolerate droughty conditions any time during the growing season. When rainfall peters out and soils hoard their water, many plants have a tough time just surviving, much less thriving. But anthocyanins can help in the same way they help juvenile leaves.
Usually drought-tolerant plants have small, thick leaves to resist water loss. Red leaves, however, can afford to grow larger because the anthocyanins help collect and maintain a healthy water reserve. Here’s where plant physiology and ornamental value happily coincide. Research on red-leaved cultivars of Viburnum and smokebush (Cotinus), for instance, has shown that these attractive choices require less watering than green-leaved cultivars. The same is probably true for many other red-leaved species whose drought tolerance has not been scientifically studied. Be prudent is using these red-leaved cultivars, however. They’re not good choices for well-watered sites, where green-leaved cultivars are more competitive.
Droughty soils can stress plants beyond just denying them water: they often contain high levels of salts. The more evaporation that occurs, the greater the concentrations of salts become. Whether they occur naturally or as a result of excessive fertilization, these salts can doom sensitive plants.
Water Crystals (Hydrogels, Dry Water)
THE PRODUCT
Water-absorbing chemicals are added to soils during transplanting or when normal irrigation isn’t possible.
THE SUPPOSED BENEFITS
Water crystals absorb large amounts of water and slowly release it into the surrounding soil as it dries.
HOW PLANTS RESPOND
Hydrogels are very good at retaining water, and they do release it slowly as plant roots take up water from the soil. But there will come a point when the gel won’t release any more water, and instead will suck it up from the surrounding soil, as well as from plant roots. So using hydrogels is not a good substitute for normal watering. Using them around new transplants, with their actively growing roots, is a bad idea. In fact, research has demonstrated that these gels aren’t any more effective than just using a decent organic mulch to help keep the soil hydrated.

Hydrogels not only provide water to roots, they can suck it away as well.
A Salty Tale
As a youngster growing up in the sometimes soggy Pacific Northwest, I was often sent outside with the salt shaker to kill slugs in my mother’s garden. (Gives a whole new meaning to “salt of the Earth,” doesn’t it?) It was with a mixture of horror and amazement that I would watch the slug shrivel before my eyes. Later I would find a miniature carcass left where a plump slug had happily munched petunias only hours before. Though I no longer use the seasoning method of slug control, the experience taught me an unforgettable lesson about how salt affects water movement.
Once more, we just need to remember the water movement rule: “Water always moves to where water isn’t.” Salt dissolves in water, so adding it to the slimy skin of a slug draws water out of the slug’s body. In much the same way, salt in the soil can pull water away from plant roots, making it more difficult for the plant’s internal irrigation system to function.
Once again, it’s anthocyanins to the rescue! Because anthocyanins help plant tissues hang onto their internal water, a tug-of-war between salt molecules and anthocyanins ensues. We can find anthocyanins in the roots, stems, and leaves of plants adapted to salty environments. Remember, it’s not just the roots that need to pull water out of the soil. The leaves need to pull water from the roots as well.

The tiny glistening dots on this ice plant (TOP) are salt storage bladders (BELOW).
Halophytes (literally salt plants) are exquisitely adapted not only to tolerate salty conditions but to revel in them. The species scientists have studied so far appear to use anthocyanins to hoard internal water. Mangrove trees, which inhabit salt marshes, collect anthocyanins in their leaves. In desert environments, ice plants such as Mesembryanthemum crystallinum collect anthocyanin-like pigments in bladders at the tips of their leaves. These little red bladders pull water from the soil, through the roots, and into the leaves, leaving the salt behind. It’s likely that other plants living in salty soils or in coastal regions have the same ability to use anthocyanins in extracting and preserving water.
A Ripe (and Red) Old Age
We’ve seen some definite advantages for young leaves and mature leaves to hoard anthocyanins, but what about old leaves? Specifically, what about all the trees and shrubs whose leaves turn red in autumn before they finally die and fall off? They’re certainly not expanding any more, and the chance of drought stress is remote. Yet every autumn both gardeners and non-gardeners alike wonder what causes leaf color change and why it varies so much between different species and different climates.
First, let’s consider what deciduous woody plants are experiencing in autumn. The days are often bright and warm, and photosynthesis chugs right along. Evenings, however, are markedly cooler and on clear nights we start getting light frosts. The foliage of deciduous species, unfortunately, isn’t able to withstand severe freezing, so trees literally cut their losses by dropping these leafy liabilities. It’s a delicate balancing act trees must perform: on one hand they need to photosynthesize as far into autumn as possible, but on the other they must mobilize leaf nutrients and shuttle them elsewhere for winter storage before killing frosts move in and leaf nutrients are left stranded in dead foliage.

Sugar maple leaves change colors with the onset of cooler autumn temperatures.
The most protected parts of trees and shrubs are the large woody tissues—branches, trunks, and roots—and this is where nutrients are tucked away until spring. It’s fairly easy for leaves to break down proteins into amino acids and starch into sugars, and these smaller building blocks are easier to transport through the plant. But individual sugar molecules, such as glucose and fructose, are reactive and have a tendency to bind to other reactive compounds inside cells. To prevent molecular mayhem from gumming up the works, leaves use carrier molecules to bind the sugar and carry it away. What are these carrier molecules? Anthocyanins, of course!
A combination of short days, cool nights, and increased sugar levels flips the switch on the anthocyanin assembly line. Newly minted anthocyanins are paired up with sugars and begin their seasonal migration, just like the snowbirds heading for points south. At the same time, other salvageable leaf structures are being broken down and transported for storage: anything that can be recycled will be. As photosynthesis grinds to a halt and sugar levels decline, the levels of anthocyanins decrease, too. Eventually, all that’s left of the leaf is a dried brown shell.
Why are anthocyanins so important in this process? In other words, why isn’t there some other carrier molecule? It turns out that anthocyanins are providing double, maybe even triple, duty in autumn leaves. They transport sugars, they help the leaves retain water, and their antioxidant activity may even protect the leaves from environmental damage from daytime sun or evening frost during this sensitive stage.
You may know how important antioxidants are to human diets. They’re also important to the plants that manufacture them. Anthocyanins are incredibly powerful in this regard; they’re even stronger than vitamin E. As leaves begin to die off, the protective cuticles erode and the internal tissues become more sensitive to sunlight and other environmental factors. The stress that these factors cause results in the formation of reactive oxygen radicals and other potent, destructive molecules that poke holes in membranes and generally cause death and destruction. Anthocyanins neutralize these radicals, allowing the leaves to stay active until nutrient transportation stops.
The coolest thing about this entire shutdown process is that we can see it happening. Let’s look at a sugar maple (Acer saccharum) and its stunning display of autumn color. First, the green leaves seem to turn bright red overnight, as the anthocyanin production machinery turns on full bore. While anthocyanins transport sugars, the pigments in the chloroplast are being disassembled for parts. The leaves lose their greenness, and we see the bright red-orange combination of anthocyanins and carotenoids appearing at the margins and working its way inward. The carotenoids disappear next, and the leaves turn yellow as the last chemical stragglers are swept out of the leaf and into their winter destination. The connective tissue between the leaf and the stem is sealed off, and the leaf dies. The stage is now set for winter’s arrival.
It’s a Cold, Cruel World
Winter in the garden can be both beautiful and desolate. Songbirds have migrated to escape potentially deadly freezes. Squirrels and other winter visitors brave the cold with thickened fur, layers of fat, and other protective methods. Plants have a variety of winter survival tactics, too. Annual flowers and weeds simply escape winter altogether, flowering and dying before temperatures turn cold and leaving their seeds behind to carry on the following spring. Hostas and other herbaceous perennials lack leaf protection against cold and die back to the root crown insulated in soil and mulch. The buds and woody tissues of trees and shrubs have chemical and physical barriers to ice invasion, but leaves do not. As we’ve seen, deciduous species shed their leaves, but what about evergreens like rhododendrons, cedars, and ivy? Not only are the leaves of these species still alive, they keep churning out sugars all winter long. They’re full of water, so why don’t they freeze solid like birdbaths and garden hoses?
Well, let’s look at those birdbaths and garden hoses first. Their water is relatively undiluted, meaning the water doesn’t have a lot of dissolved sugars or other chemicals in it. When ice first starts forming, it begins a chain reaction: wherever ice touches liquid water, that water is transformed into ice. Ice formation is slow at warmer temperatures, say just around freezing, but as the temperature drops, freezing is faster. (The freezing rate is important when it comes to predicting plant damage, and we’ll come back to this topic in a bit.)
First we’re going to do an experiment, one that is easy enough for you to do at home. We’ll start with two very clean, narrow glass containers like test tubes. After filling them about halfway with water, we brush a little dust into one of the tubes and put them both in a freezer. After an hour or so, we check on them to see if they’ve begun to freeze. The one we sprinkled with dust is starting to get an icy film on the surface, but the other is still in liquid form. We check on the first one in another hour and can see ice has grown along the inside surface of the tube, but there’s still a core of liquid water left in the middle. The other test tube is still unfrozen. Finally, after three hours the first tube has filled with ice, but the second is still liquid. The temperature of the water in this second tube is actually below freezing, maybe around 28°F. This is known as supercooled water.
Now the fun begins. We take a toothpick and touch the surface of the water in the second tube. Immediately the entire tube freezes solid, so it looks just like the first tube. The supercooled water was waiting for a nucleator—in this case the toothpick—to start the chain reaction. We’ve just observed the differences between slow and fast freezing events
Before we move on, let’s consider why we only filled those tubes halfway up. You know what would have happened if they were brimming with water: the ice would have expanded over the edge and made a mess. (Water is unusual in that its solid form has lower density than its liquid form and therefore takes up more space.) If you’re as absent-minded as I can be, you too may have put a can of pop or bottle of wine in the freezer to cool it quickly and then forgotten about it until much later. You know what you’re going to find when you finally open the freezer again!
Now, let’s substitute plants for the test tubes and see what happens. When water freezes quickly inside of plant cells, the expanding ice stretches the cell membranes until they split open. When you place lettuce or other greens too close to the cooling elements in a refrigerator you’ll see the results: soggy, glassy-looking leaves. The same can happen in your garden, though it’s rare for temperatures to decrease that rapidly. Instead, what usually happens is that ice forms slowly in the leaves, particularly in the air spaces outside of the cell walls. This intercellular space contains water and is surrounded by nonliving tissue, so when the water freezes it doesn’t cause any damage. So far, so good.
Remember, “Water always moves to where water isn’t.” As far as the plant is concerned, ice isn’t water. Ice can’t be taken up by roots or dissolve nutrients. The intercellular spaces become drier as ice forms, sucking water away from the nearby cells. Now the living cells in the leaf are dehydrating and becoming water stressed. The longer the freezing in the air spaces continues, the more dehydrated the living cells become. If it goes on too long, the cells eventually die. But they die from lack of water, not from ice damage.
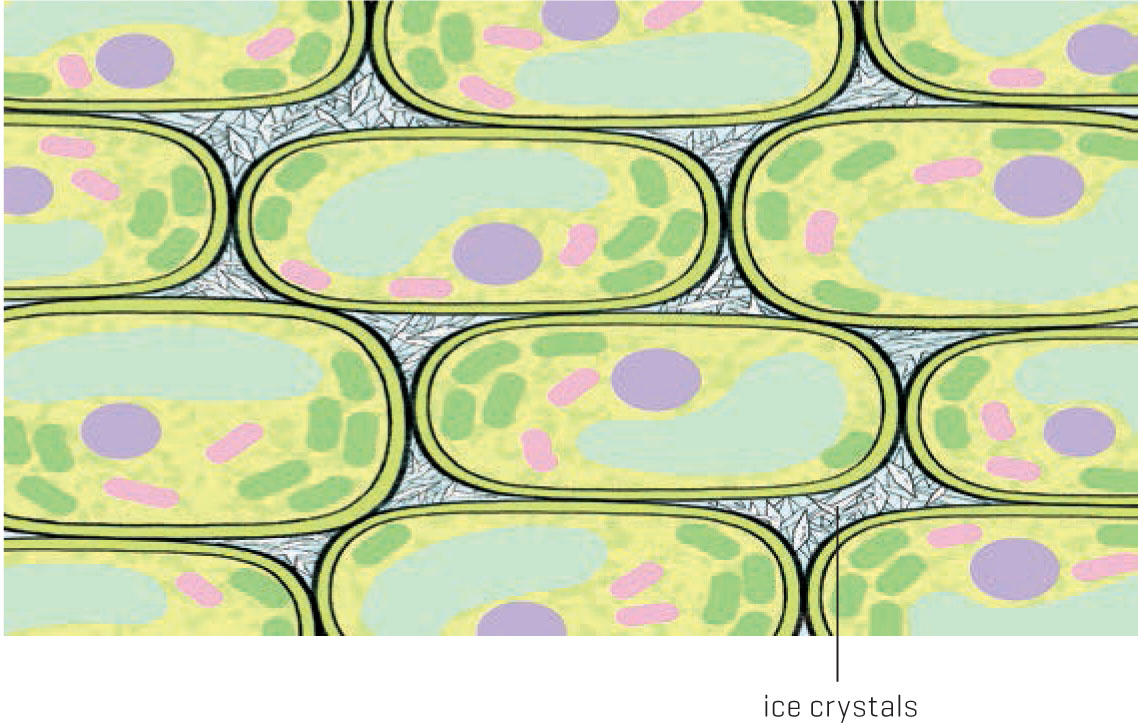
Plant cells contain dissolved substances that keep cellular water from freezing, encouraging ice to form between the cells instead.
How do evergreen leaves combat this frozen doom? You know the answer by now: it’s anthocyanins! Low temperatures cause leaves to accumulate anthocyanins in a wide variety of garden favorites, both evergreen and deciduous, including maples (Acer species), red-twig dogwood (Cornus sericea), smokebush (Cotinus), Euonymus species, English ivy (Hedera helix), Oregon grape (Mahonia repens), apple (Malus domestica), pines (Pinus species), peach (Prunus persica), and sumac (Rhus species). We’ve already seen how anthocyanins help deciduous leaves prepare for their autumnal demise, but evergreen leaves accumulate anthocyanins as a kind of antifreeze. Once accumulated, anthocyanins help red-tinged leaves of Oregon grape, viburnum, and rhododendron hang onto their cellular water better than green leaves of the same plant. Water that’s bound inside the cells is not able to migrate easily into intercellular spaces, so the leaves are able to withstand colder temperatures. They can accumulate so many anthocyanins that their leaves are visibly redder than they were during the summer. When spring returns along with warmer temperatures, winter-reddened leaves regain their usual green color. This doesn’t happen in all evergreen species, but those that do turn red sure brighten up our winter landscapes.
This freeze-avoiding benefit that anthocyanins offer is also useful in those juvenile leaves we talked about earlier. Leaf buds are tough little structures, resistant to hard winter freezes, but once they break dormancy the emerging leaves are susceptible to late spring frosts—even mild ones. Ice forms quickly on top of and inside these tissues, and that’s the end of those leaves. But anthocyanins can lower the freezing point of water, just like adding salt to sidewalks keeps them from freezing. Juvenile anthocyanins allow leaves to survive their youth, much like immunizations help our children avoid fatal diseases and become teenagers. (And we have the pleasures of gardening to help us survive teenagers.)
Anthocyanins and Heavy Metals
I’m continually amazed at how anthocyanins can protect plants from so many different environmental insults. And few environmental insults can have as severe consequences as those of heavy metal contamination.
Heavy metals are a loose collection of mostly metallic elements with significant effects on life. Some heavy metals, such as iron, are nutritional requirements of animals and plants alike: we can’t live without them (at least in small quantities). Others have no known benefit and in large enough quantities can be toxic. Many gardeners are well aware of this problem, testing their soils for lead, arsenic, and other common heavy metals before planting vegetables and other edibles.

The red leaves of Oregon grape are more cold hardy than green leaves.
One of the dangerous things about heavy metals is that, because they are elements, they don’t break down to nontoxic forms. Worse, they can be accumulated over time. We’re told, for instance, to limit our consumption of swordfish because it can be high in mercury. So, the more swordfish we eat, the more mercury we accumulate. In high enough doses, mercury and other toxic heavy metals start to interfere with our enzyme activity and eventually can be fatal. Therefore, we try to limit our exposure to these metals as best we can, such as getting rid of lead in gasoline and paint and removing arsenic from wood preservatives and pesticides.
Because plants are stuck wherever it is they’re growing, they adapted to heavy metals long ago. For some species, this means not growing where toxic heavy metals exist. But other plants tolerate and sometimes concentrate these elements, and because they have few competitors they grow profusely. There are arsenic-accumulating ferns, mercury-mining mustards, and cadmium-collecting crops. They employ different survival strategies, sometimes avoiding metal uptake altogether and sometimes taking it up but isolating it somewhere far away from sensitive enzymes. We can think of this last strategy as putting toxic heavy metals in time out.
Part of keeping heavy metals locked away is to make them less toxic to the cells, and this is where anthocyanins come in. Anthocyanins bind to reactive compounds such as sugar, and heavy metals are also reactive. Researchers have discovered plant species that naturally take up and bind heavy metals to anthocyanins, detoxifying the metal and allowing the plants to survive on these hostile soils. The flowers of one such plant, Linanthus parviflorus (a member of the pink family), turn from white to pink when exposed to soils containing heavy metals! It’s like a botanical soil testing system.
A Watery Conundrum
We’re going to step into my front yard and look at a Korean dogwood (Cornus kousa) that I planted eight years ago. This lovely tree was my poster child touting the benefits of wood chip mulches. But an odd thing happened after the first five years: the leaves became progressively smaller, redder, and less numerous than previous years. Finally, we realized that the tree was going to die unless we took drastic steps. When we dug it up, we found a perched water table (thanks to the house builders who buried clay excavated from the foundation 30 years ago) and a root system that had rotted away to nearly nothing. Why does waterlogged soil turn leaves red?
The pore spaces in garden soil, the areas between soil particles, are filled with either liquid (usually water) or gases (usually carbon dioxide and oxygen). So any time there’s standing water in your garden soil, you can bet there’s little oxygen. This oxygen deficit, or hypoxia, can doom plants that aren’t adapted to wet soils, like my Korean dogwood.
What causes hypoxia? Sometimes it’s flooding or a perched water table, created when soils of different types are layered onto each other. Either situation will fill those pore spaces with water. Compaction from vehicles, foot traffic, heavy equipment, and even pets can squeeze the oxygen (and the life) out of soil. Heavy clay soils, with their fine particulate texture, have correspondingly tiny pores and little space for oxygen.
Aeration Tubes
THE PRODUCT
Plastic pipes installed vertically into tree planting holes.
THE SUPPOSED BENEFITS
Pipes bring oxygen down to roots like a snorkel.
HOW PLANTS RESPOND
In unobstructed landscapes, tree root establishment is primarily horizontal, as surface soils are most likely to contain substantial oxygen, water, and nutrients. In an enclosed area such as a container, roots contact the wall and grow deeper into the soil. An aeration tube may work in such a small volume of soil. However, a single pipe is unlikely to provide enough oxygen in the landscape, and field research has not supported the use of aeration tubes in outdoor settings.

Root snorkels aren’t effective in aerating soil or enhancing tree survival.
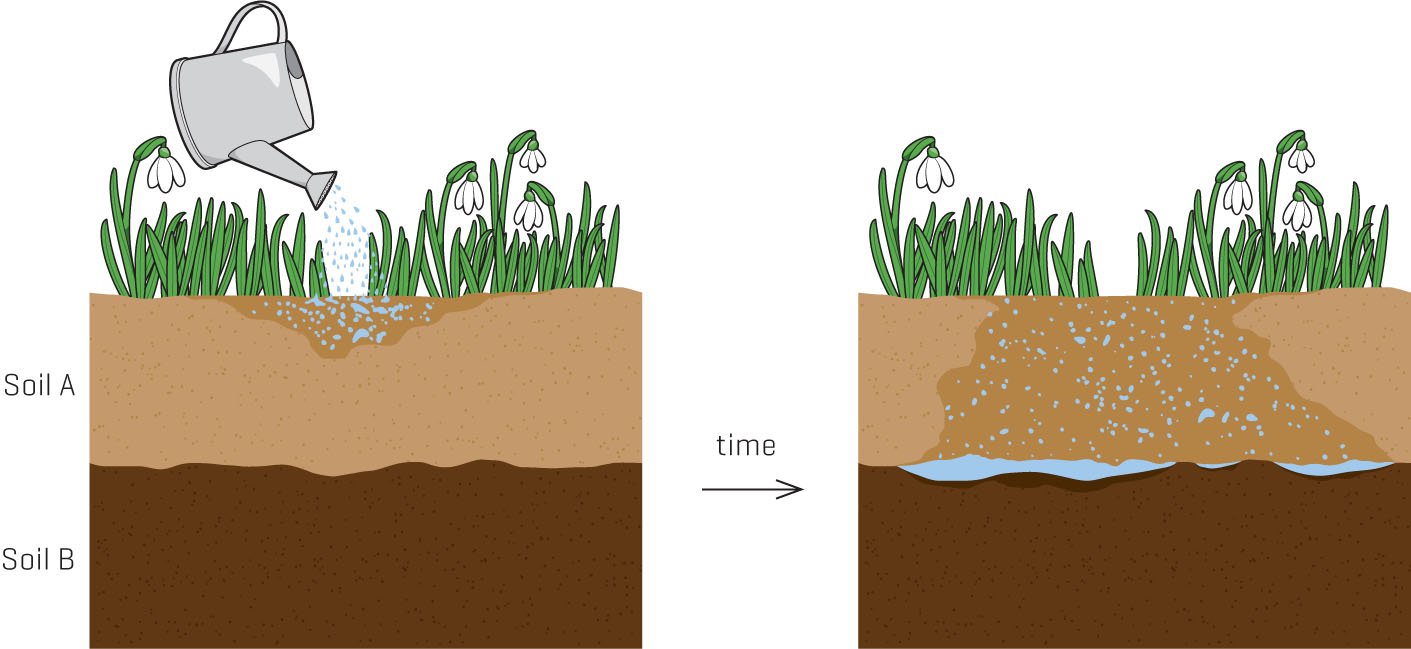
Contact between different soil types restricts water movement, leading to a perched water table.
Now let’s look at my dogwood’s pathetic root system. All but one of the woody roots had rotted away, and the remaining system consisted of fine roots near the soil surface where oxygen could still be found. This shrunken root system could hardly support the substantial crown that developed in the first five years. But unlike the situation in most drought-stressed trees, the leaves on my waterlogged tree didn’t die or even become brown. They were just small and red.
Our reliable, multipurpose anthocyanins were on the job again. Even though water uptake by the roots was a fraction of what it should have been, anthocyanins prevented evaporation from the leaves. Compared to soils with plenty of available oxygen, researchers have documented increased anthocyanin production in the leaves of trees in hypoxic soils, including apple (Malus species), pear (Pyrus species), madrone (Arbutus menziesii), and red maple (Acer rubrum). Generally, once the problem is solved (the floods recede or you transplant your unhappy plants), the leaves turn green again as the anthocyanins are broken down. New leaves are bigger and healthier with improved root water uptake. And that’s exactly what happened to our Korean dogwood when we moved it to better-drained soil.

The bulls’ eye pattern on these salal leaves is due to fungal infection.
Pushing the Red Button
Science has only grazed the tip of the anthocyanin iceberg. It’s likely that these leaf pigments are instrumental in protecting plants against a slew of other environmental insults. For instance, leaves may turn red when they’re short of certain nutrients, such as boron, magnesium, nitrogen, phosphorus, sulfur, and zinc. It’s unclear what anthocyanins are doing in these situations, but because so many nutrients are associated with controlling water levels in the plant, anthocyanins may reduce water loss when these nutrients are deficient.
When soil pH is lowered beyond what’s normal for plants in that area, red leaves may develop in response. Because acidic pH makes heavy metals more available for root uptake, it’s likely the anthocyanins are binding those metals and storing them away from sensitive enzymes.
Finally, what gardener isn’t intimately familiar with the bright red bulls’ eye pattern caused by fungal disease on leaves? The center is the original point of infection, which dies and turns brown as the fungus invades the leaf. The reddest part of the pattern is on the battlefront between healthy leaf and fungal invaders. Are the anthocyanins actively fighting the fungus, perhaps using their antioxidant powers? Or are they just helping reduce water loss as the leaf tissues are exposed to the elements? Because we may also see this reddening in response to bacterial attack or other wounding, it may be an attempt to reduce dehydration along those damaged edges.
I guess I’ve exposed my plant biochemistry geekiness in spending an entire chapter showcasing anthocyanins. We know that they can tell us about soil conditions, water availability, and even the changing of seasons. And I firmly believe that science will continue to demonstrate that these pigments are major players in many plant defensive moves. Savvy gardeners can use this easy visual cue to figure out what environmental factor plants might be responding to, and what—if anything—they should do about it.
As you might guess by now, I think anthocyanins are the coolest thing in the plant pigment world. They can tell us about soil conditions, water availability, and even the changing of seasons. But autumn reds are a response to winter’s approach. They aren’t part of a plant’s internal clock. So how do our deciduous trees know when autumn is in the air?