How Plants Work (2015)
4 Transforming Sunlight into Sugar
IT’S MIDSUMMER. The cool, wet weather of spring is a distant memory. Insects whir and buzz, and the garden overflows with plump leaves and flowers. But it’s a different story when we walk over to the lawn. To conserve water we try to limit irrigation, but the thirsty nature of grasses means that they look a little sad and defeated in the midday sun. The exception are those annoying lawn weeds, such as crabgrasses (Digitaria species), purslane (Portulaca oleracea), and nutsedges (Cyperus species). They seem to defy drought and heat, looking like little green islands in a sea of yellow and brown. How can these weeds tolerate the dog days of summer, while our desirable grasses literally go to seed?
The answer lies within a plant’s biochemistry, specifically, its photosynthetic pathway. Gardeners know quite well that plants are solar powered: they use the sun’s energy to drive a complex set of chemical reactions that transforms a common gas (carbon dioxide) into a solid form (carbohydrates). Every time you enjoy a salad from your garden-fresh greens or lick the juice from the season’s first strawberries from your fingers, you are experiencing the delightful results of photosynthesis. It is, without a doubt, the most important biochemical reaction for life on Earth.

The green weeds in this brown lawn have a unique biochemical pathway that helps them survive hot, dry conditions.
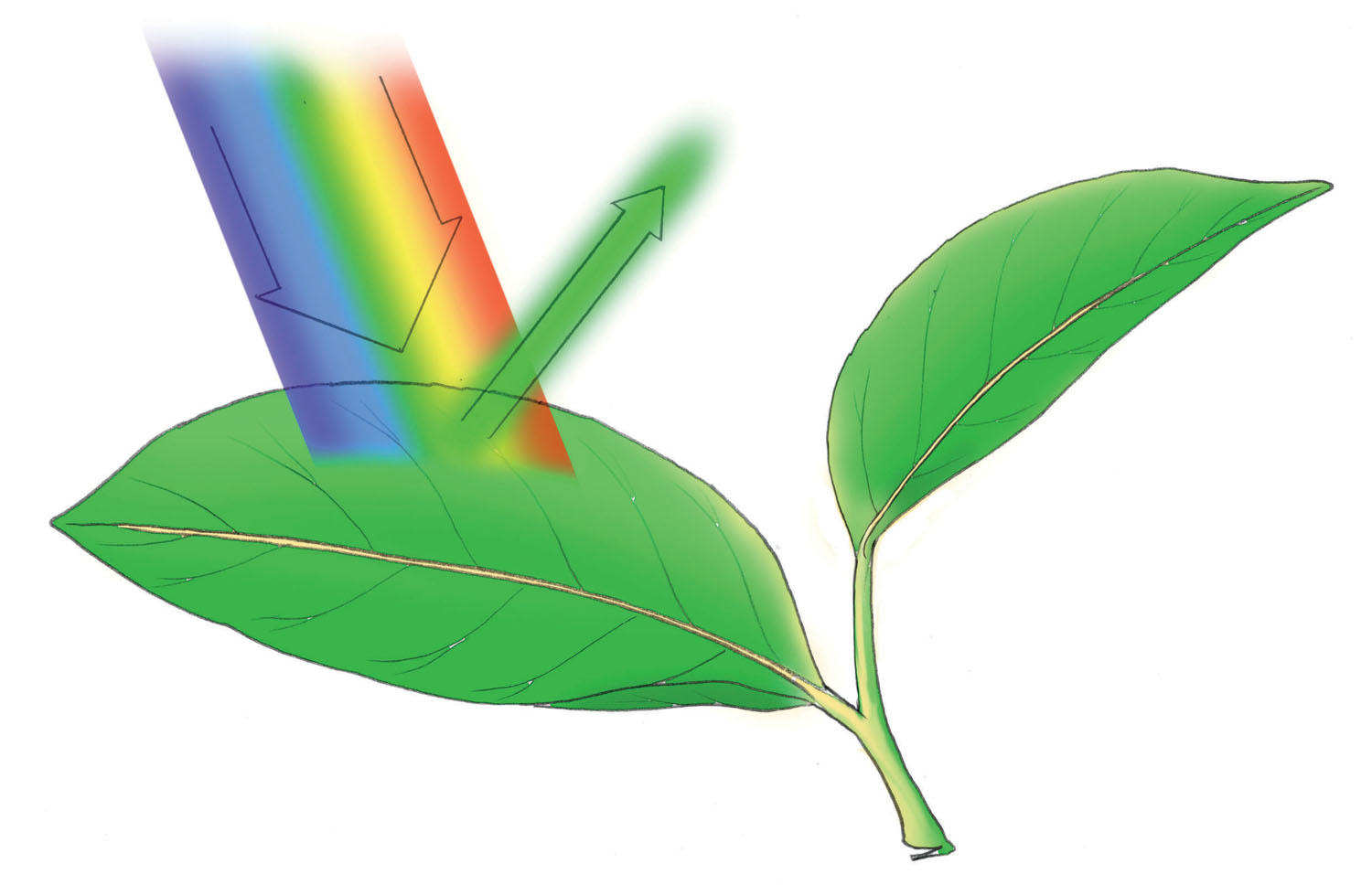
When sunlight, with its full spectrum of colors, strikes a green leaf, all the colors are captured in the leaf pigments except for most of the green light, which is reflected.
Harnessing the Sun
The first sunny day of spring is a tonic to plants and gardeners alike. Tomato seedlings in the greenhouse and potted bulbs overwintering in cold frames respond eagerly to the warmth as well as the biochemical kick-start that energy from sunlight supplies. In fact, sunlight consists of a wide range of energy including powerful, short-wavelength radiation (including X-rays) as well as weaker, long-wavelength radiation (such as radio waves). Within this broad spectrum is the narrow rainbow of visible light used to run many biological reactions, including photosynthesis. Sunlight that touches a leaf can be trapped (absorbed), be bounced back (reflected), or pass through untouched (transmitted). Only the absorbed light can be harnessed and put to work inside the plant. And for light to be absorbed by a leaf, it has to be captured by a pigment.
Two groups of pigments are used to trap light for photosynthesis: the green chlorophylls and the orange carotenoids. It surprises many people to discover that the color of a pigment is actually the result of the light that isn’t absorbed, but instead is reflected and transmitted. So chlorophyll doesn’t absorb green light well at all, but it is very good at nabbing the red and blue wavelengths. Likewise, carotenoids capture blue wavelengths but reflect the red and orange portions of the spectrum. The color you see is whatever color is not absorbed.
Let’s look at the rainbow riot of colors in my own garden in this scientific context. The red spring tulips and summer daylilies absorb every color except red, which is reflected back to our eyes, and we see red. Likewise, the yellow daffodils, green hellebores, blue hydrangeas, and purple clematis absorb all colors except for the ones that we can see. And those pure white mock orange blossoms? They don’t absorb any visible light, so we see all that white sunlight reflected back to us. A pure black flower would swallow every bit of visible light like a black hole, leaving nothing for us to see. (In reality, most black flowers have some tinge of red, blue, or purple.)
Creating Sugar out of Thin Air
Now back to our green leaves, busily trapping bits of sunlight (particles known as photons). Once a photon has been captured by a photosynthetic pigment, it’s used as a biochemical booster in creating plant food. The most important step of this long and complicated process requires leaves to suck carbon dioxide from the atmosphere and attach it to a small, five-carbon organic compound. This creates a six-carbon compound that immediately splits into two three-carbon compounds, which are biochemically poked and prodded until they end up in a sugar molecule. The term carbon fixation is used to describe this transformation of carbon from a gas to a solid form. All life on Earth depends on this reaction. (Well, except for anaerobic microbes whose dependence on other biochemical pathways has the unfortunate distinction of producing hydrogen sulfide and other Eau de Swamp gases.)
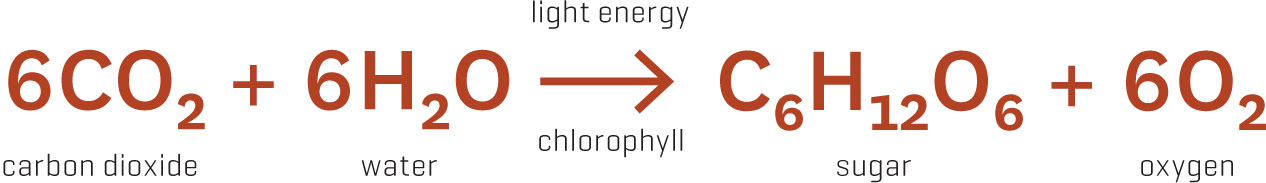
Plants use the sun’s energy to transform carbon dioxide and water into sugar and oxygen.
If photosynthesis is the most important biochemical process on Earth, then the enzyme that glues carbon dioxide onto that five-carbon compound is the most important enzyme. It has an important-sounding name as well: ribulose 1,5-bisphosphate carboxylase oxygenase. Fortunately, plant physiologists love acronyms, and I’ll call this enzyme RuBisCO. (Fun fact: RuBisCO makes up 30 to 50 percent of the total protein in leaves, making it the most common protein on a global scale. It’s the letter E in the biochemical Scrabble game.)
RuBisCO is an ancient enzyme. The single-celled organisms that contained it evolved long before dinosaurs or cockroaches or even earthworms appeared on Earth. It’s found in every photosynthetic organism, including bacteria and algae as well as higher plants. Earth was quite a different place when RuBisCO showed up, primarily because there was no free oxygen in the atmosphere. Life consisted of primitive microbes existing in oxygen-free (anaerobic) environments, like those that thrive in wet, poorly drained soils today. In much the same way, RuBisCO works best under anaerobic conditions, meaning it’s able to do its job gluing carbon units effectively.
Oxygen: A Toxic Byproduct of Photosynthesis That We Can’t Live Without
There’s an important chemical reaction that occurs when leaves absorb photons for photosynthesis, and that’s the splitting of water molecules. When water molecules are split, they release the electrons needed to drive photosynthesis and they also form molecular oxygen. Primitive Earth had no oxygen gas floating around, and this new chemical was a lethal, corrosive pollutant. As photosynthetic organisms evolved and oxygen began to accumulate in the atmosphere, a catastrophic extinction of early life forms followed. (Most modern anaerobes are now confined to underwater or underground habitats, where oxygen levels are low enough for them to survive.) New life forms appeared on Earth that could tolerate oxygen, and now most life on Earth is utterly dependent on a gas that was once a deadly pollutant.
Like most modern-day terrestrial organisms, plants not only adapted to the presence of oxygen but now use oxygen to break down carbohydrates and other organic compounds more efficiently. RuBisCO, however, continues to pose a problem for many plants because of its sensitivity to oxygen. Under certain conditions, this sensitivity can cause plants to die back—or just plain die.
Photorespiration: The Ultimate Weight Loss Program for Plants
Unfortunately, RuBisCO is rather sloppy in its ability to bind carbon dioxide for eventual attachment to that five-carbon molecule. When oxygen is around, RuBisCO preferentially binds it instead of carbon dioxide. (Kind of like choosing triple chocolate decadence over fruit on the dessert tray, even though the fruit’s better for you.) Therefore, RuBisCO works most efficiently when leaf carbon dioxide levels are high and oxygen levels are low, temperatures are moderate, and water is plentiful. But botanical Fantasyland and reality don’t always coincide.
When RuBisCO hooks up with oxygen instead of carbon dioxide, the plant’s biochemistry shifts from photosynthesis to photorespiration, in which photosynthetic cells consume chemical energy and oxygen, producing carbon dioxide. Plants experience several negative outcomes when oxygen throws a monkey wrench into the photosynthetic machinery.
When oxygen is glued onto the five-carbon compound instead of carbon dioxide, a different five-carbon compound is created that can’t be used in forming sugars. This deceptively simple chemical process causes several serious problems for the plant. First, RuBisCO is temporarily taken out of commission when it binds oxygen, meaning it can’t bind carbon dioxide. Low levels of available RuBisCO mean slower photosynthesis. Second, the five-carbon compound that’s usually bound to carbon dioxide has been wasted and has to be regenerated. Third, the regeneration process for making the five-carbon compound includes a step that releases carbon dioxide (the respiration part of photorespiration). Finally, all of these processes require energy to repair the problems caused by that single step of binding oxygen.
Unlike regular respiration, which all living organisms use as a way to create energy currency from stored food supplies, photorespiration uses up energy and generates useless chemicals that have to be reformulated. This is such a hugely negative process that plants in hot, bright, and/or dry environments can lose nearly half of their newly formed carbohydrates, which means they are losing weight. Think about struggling urban street trees in the middle of summer. Is it any wonder that many never seem to thrive?
Tweaking the System: C3 and C4 Plants
Then how do plants survive in environments that aren’t cool, moist, and rich in carbon dioxide? Primitive plants were restricted to aquatic environments and forests, where photorespiration wouldn’t pose much of a problem. In more hostile conditions, plants continually modify their biochemistry and sometimes their appearance, because they can’t just get up and leave for greener pastures. Some plants crank out additional RuBisCO to swamp the system, whereas others tweak this enzyme to be more particular in choosing carbon dioxide over oxygen. By far the most interesting plants are those that dramatically overhaul basic photosynthesis and figuratively thumb their noses at photorespiration.
To keep things as simple as possible, I’ll refer to basic photosynthesis as the C3 pathway because carbon dioxide is used to form two three-carbon compounds. All plants follow the regular C3 pathway, but some have developed extra biochemical steps to avoid photorespiration. The first of these steps binds carbon dioxide to a three-carbon compound, forming a four-carbon product, so this series of extra steps is called C4 photosynthesis. To fix carbon, the C4 pathway uses a completely different enzyme, phosphoenolpyruvate carboxylase, which I’ll mercifully refer to as PEPcase. Unlike RuBisCO, PEPcase is strongly attracted to carbon dioxide and binds it tightly instead of oxygen, so the C4 pathway isn’t affected by photorespiration. Unfortunately, the four-carbon compound can’t be used in C3 photosynthesis, and eventually a molecule of carbon dioxide is clipped off deep inside the leaf.
Now if you’ve been keeping track of all these biochemical contortions, you might well wonder how these pre-C3 biochemical steps do a plant any good: it’s just spent chemical energy fixing and then unfixing carbon dioxide. Where’s the benefit?
A CARBON DIOXIDE MOLECULE SAVED IS A CARBON DIOXIDE MOLECULE EARNED
Let’s look at some of the plants that have evolved C4 photosynthesis. They include more than half of all grass species as well as some herbaceous annuals and perennials, but no conifers. If you could crawl into a stomate on a giant C4 leaf and wander over to a vein, you would see it surrounded by a ring of tightly packed cells, known as the bundle sheath cells. You’d have a tough time breathing here, because that tight packing of cells means very few air spaces and very little oxygen or any other gases for that matter. The low-oxygen environment bounded by the ring of cells is where RuBisCO is conveniently stored (remember, RuBisCO just can’t resist oxygen). These rings are the final destination of the four-carbon armored cars, which release carbon dioxide once inside. So C4 plants have developed safe houses where RuBisCO can bind carbon dioxide without interference from oxygen, stymieing theft by photorespiration.
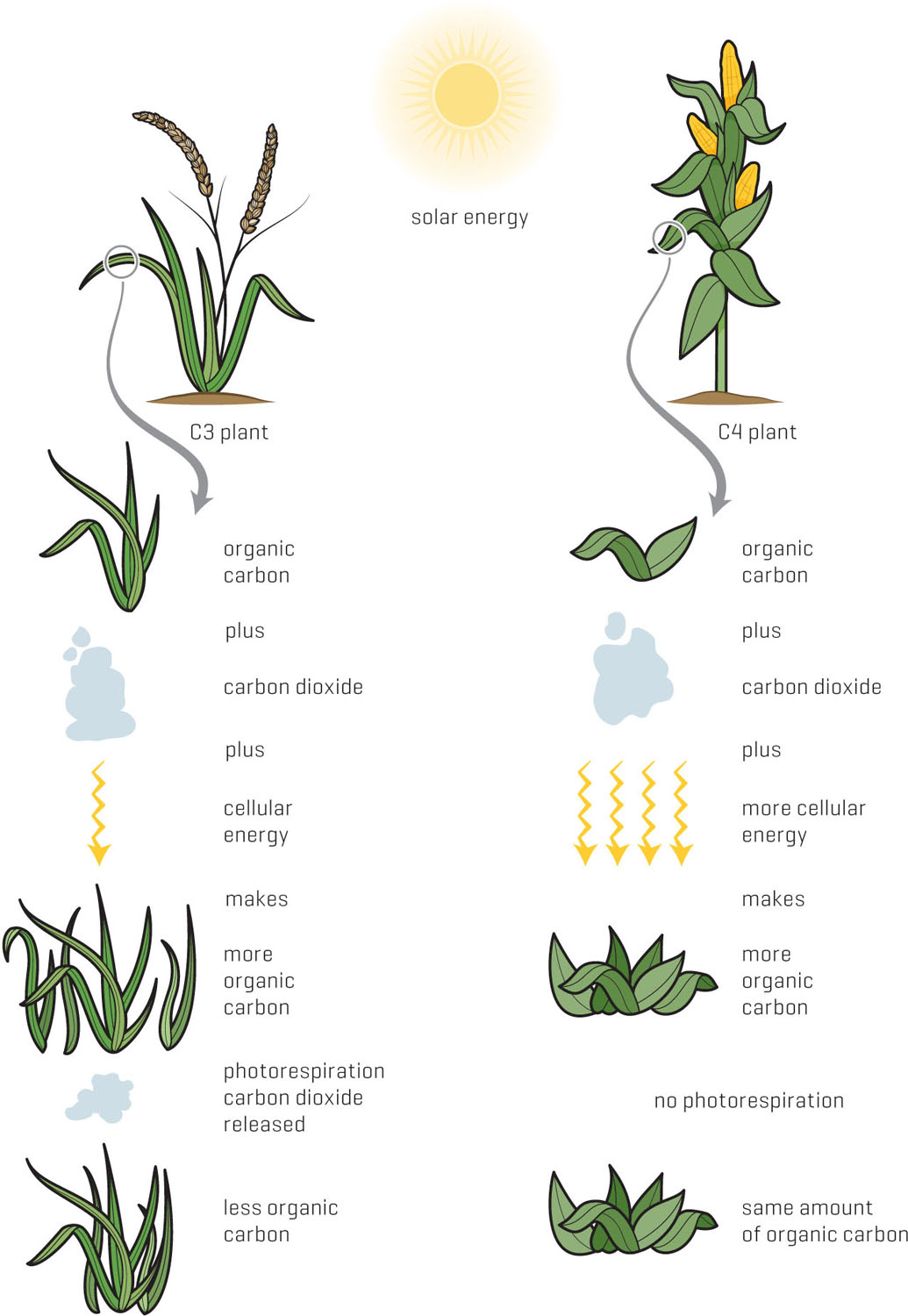
C3 plants use less cellular energy to fix carbon, but they lose carbon to photorespiration if conditions are not favorable. C4 plants use more cellular energy to fix carbon, but they don’t lose it once it’s fixed, meaning they continue to grow in conditions unfavorable for C3 plants.
The biochemical brilliance of the C4 pathway is that carbon dioxide can be fixed into a solid form in hot, dry, and bright environments without interference from photorespiration. The four-carbon compounds are like armored cars shuttling precious carbon cargo to various parts of the leaf before releasing it again as carbon dioxide. The ultimate function of C4 photosynthesis is to accumulate carbon dioxide in oxygen-free tissues.
However, the cellular energy needed to run this carbon dioxide armored transport service is significant. It takes almost twice as much energy to make sugar by C4 photosynthesis as compared to the C3 pathway. In warm environments like tropical grasslands, the energy costs are minimal compared to the advantages of avoiding photorespiration. RuBisCO is protected from oxygen and binds carbon dioxide at maximum speed. C4 plants save energy by not manufacturing extra RuBisCO and by not having to remake the five-carbon compounds wasted in photorespiration. Finally, when the environment is hot, bright, and/or dry, the net gain of carbon dioxide fixed outweighs the energy costs of the additional C4 steps.
WHY C4 WEEDS ALWAYS WIN IN YOUR DRY SUMMER LAWN
Let’s look at that dry summer lawn again. Bluegrass, fescue, and other cool-season C3 grasses that do so well in the spring (and in moderate climates in the winter) have gone dormant. The roots are still alive, but the blades have died back. Once day temperatures get much above 80°F these species take a break until cool moist conditions return. The C4 plants—the maddeningly healthy weeds we saw earlier—are in their element, photosynthesizing briskly with little interference from the dormant species. When temperatures drop and rains return, the extra photosynthetic steps of C4 plants become an energy liability instead of an asset, and the C3 plants return in full force.
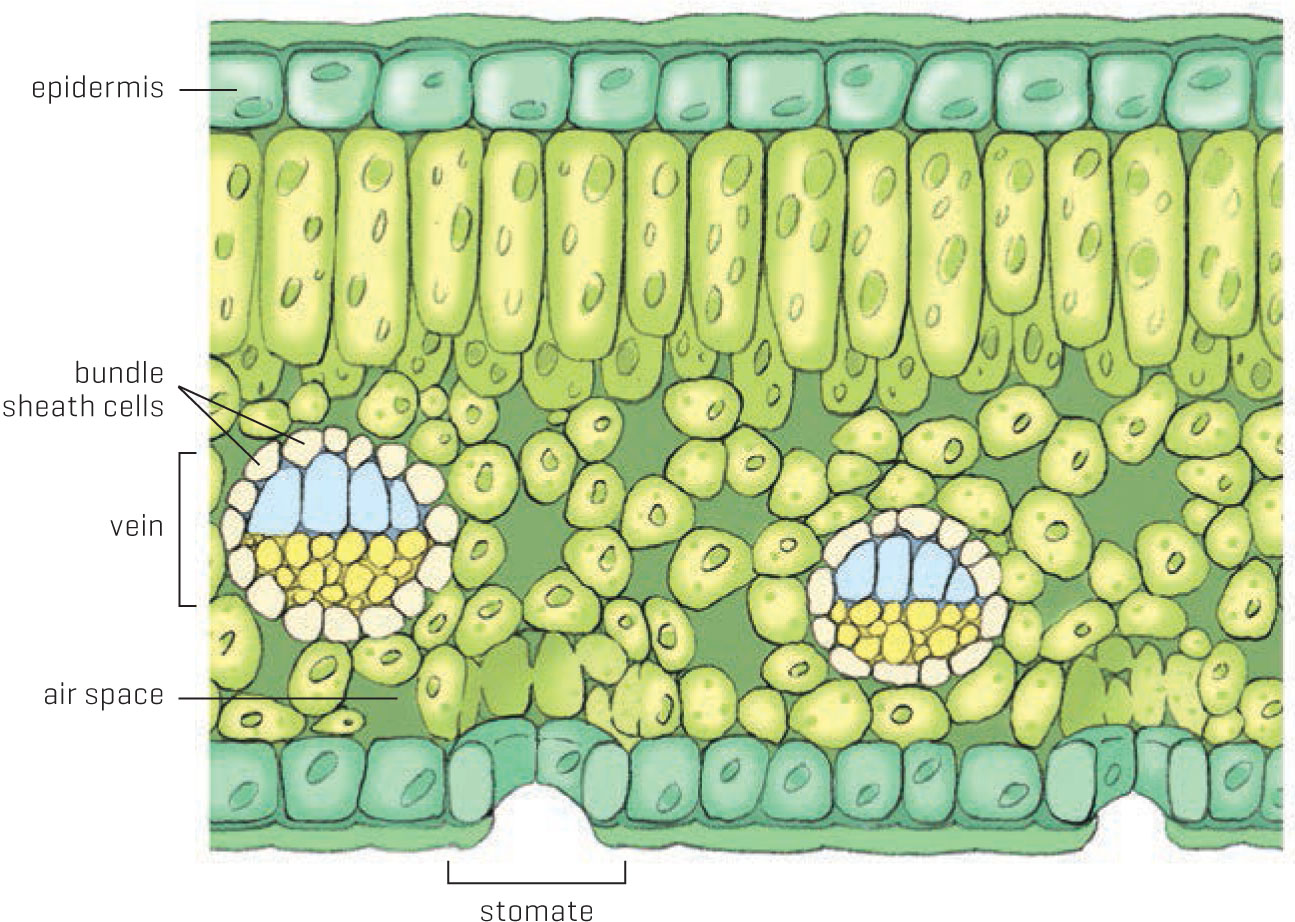
The interior leaf architecture of C4 plants allows them to concentrate carbon dioxide for photosynthesis.
As tenacious as crabgrass and other summer lawn weeds can be, even they can’t survive extended summer drought. Though their photosynthetic mechanism is still functional, like most garden plants they are vulnerable to dehydration. Without some minimal water input, they too will go dormant.
Variation on a Photosynthetic Theme
My neighbor has a green roof and she never waters it, even in the summer. Peering over the gutter, I can see it’s abundantly planted with different stonecrops (Sedum species) and hen-and-chicks (Sempervivum species). There’s hardly any soil, definitely no shade, and yet it’s a virtual ocean of succulent greenery. Surely these can’t be C3 plants, and they don’t look like any of the C4 weed species in the lawn. How do these little succulents survive and thrive in this environment?
A Baker’s Dirty Dozen
Weeds that are C4 species can make a gardener’s life miserable. Now you know why they do so well in the summer.
✵ Barnyard grass (Echinochloa crus-galli)
✵ Bermudagrass (Cynodon species)
✵ Black medic (Medicago lupulina)
✵ Cogon grass (Imperata cylindrica)
✵ Crabgrass (Digitaria species)
✵ Foxtail (Setaria pumila)
✵ Goosegrass (Eleusine indica)
✵ Johnsongrass (Sorghum halepense)
✵ Nimblewill (Muhlenbergia schreberi)
✵ Nutsedge (Cyperus species)
✵ Pigweed (Amaranthus species)
✵ Purslane (Portulaca oleracea)
✵ Spurge (Euphorbia species)
While the C4 photosynthetic pathway deftly sidesteps photorespiration, it gets hammered in perpetually arid environments. Leaf stomata must be open for carbon dioxide to enter, but then water vapor escapes. Shutting the stomata slows photosynthesis, and the C4 pathway becomes less efficient. Deserts, environments that receive less than 10 inches of rain per year, are characterized by succulent species in the cacti and euphorb families. First and foremost, desert plants must be able to survive drought. To do this they store water in fleshy tissues and reduce water loss by closing their stomata during the day. So, our little stonecrops collect carbon dioxide only at night when the stomata are open. But photosynthesis requires sunlight. How can biochemistry be reconciled with reality? Let’s explore another fascinating side road off the photosynthetic pathway.
Gardeners and scientists alike have to marvel at how plants adapt to seemingly impossible conditions. Once again, the C4 pathway runs the carbon dioxide catch-and-release program, but this time it happens at night. (The PEPcase enzyme functions any time, day or night, and that works well for those green roof succulents.) Stomata open wide as the sun sets, allowing carbon dioxide to flow in as oxygen from photosynthesis flows out. The biochemistry in our succulents is exactly the same as for C4 plants: PEPcase binds carbon dioxide and creates the four-carbon compounds. But in succulent species these carbon dioxide carriers remain in the same cells as where they’re produced, parked overnight in large, water-filled storage structures called vacuoles. At sunrise, the stomata close down, the carbon dioxide carriers pull out of their overnight parking lot, carbon dioxide is unloaded into the same cells where it was picked up a few hours earlier, and RuBisCO begins the C3 pathway in a moist, warm, carbon dioxide-rich environment.
Here’s an interesting tidbit about the four-carbon compounds in C4 photosynthesis: they are all organic acids. This helps explain early observations on the Crassulaceae (meaning fat plant), the family that includes stonecrops and hen-and-chicks, which described them as having a “daily acid taste” in the early hours—kind of a cactus version of morning breath. It’s the acid-storing property of succulents that gives this photosynthetic detour its name: Crassulacean acid metabolism or CAM.

Well-watered euphorbs grown indoors often have leaves (LEFT), whereas those in more natural environments do not (RIGHT).
The one drawback to CAM photosynthesis is that the stomatal shutdown at sunrise leads to an internal oxygen buildup, so photorespiration increases as the day wears on. Therefore, CAM is inefficient and expensive compared to C3 and C4, and as a result CAM plants grow very slowly in their native habitats. Yet this pathway is widespread, found in thousands of plant species belonging to dozens of plant families including the agaves, cacti, euphorbs, lilies, and orchids. What unites all CAM plants is that their native environments all experience limited water or daytime carbon dioxide at some time during the year. CAM plants enjoy growth spurts during the rainy season or when irrigated in landscapes, greenhouses, and indoors. That’s why my elkhorn euphorb (Euphorbia lactea ‘Cristata’) only took fifteen years to reach 2 feet in our house, whereas it might take many decades to reach that height in nature. Abundant water plumps succulent tissues and can even encourage temporary leaves to pop out, only to die back when water becomes scarce.
Xeriscaping
THE PRACTICE
Using drought-tolerant species (xerophytes) for landscaping in arid climates.
THE SUPPOSED BENEFITS
Drought-tolerant species don’t use much water, so homeowners will save money on irrigation.
HOW PLANTS RESPOND
Xerophytes are fascinating plants. They survive quite nicely with little water, but they are fierce competitors for water when it’s available. In response to extra water (what we call luxury consumption), they reward gardeners with a flush of leaves or a burst of flowers. But once the water is gone, they often drop some or all of their leaves and go dormant. This is typical for plants in stressful environments. However, gardeners don’t like this and will usually water the plants again to get that aesthetically pleasing response. You can end up using as much or more water keeping xerophytic plants lush-looking than it takes for drought-intolerant species. So by all means use xerophytic species, but realize you are creating a landscape that is naturally slow growing—unless, of course, you add lots of water.

Drought-tolerant plants are often water hogs.
Harnessing the Sun a Bit Too Well
Descending from my neighbor’s green roof with a newfound appreciation for the tiny stonecrop, we spot a problem across the street. Last week the exceptionally neat homeowner in residence sheared his boxwoods into regulation shapes and some of the remaining leaves are unusually pale. Closer inspection reveals they’re actually white. There are no signs of insects or disease. What could have happened to turn leaves white in the middle of summer?
Plants that love full sun can be pretty sloppy about collecting light. In fact, leaves exposed to full sun are generally small, thick, and have protective layers of wax or fuzzy hairs to prevent internal solar overload. My sun-loving garden herbs, lavender and rosemary, are good examples, with their thick needle-like leaves and waxy sheens. Or my sage plant, with its thick, woolly, lambs-ear foliage. In contrast, shade-loving plants like my giant hostas need to capture every stray photon to keep their photosynthetic machinery running. Their leaves are large and thin, with little surface protection. Species that tolerate both sun and shade develop leaves that adapt to whichever environment they are grown in. This last strategy mirrors the way that leaves develop in shrubs and trees: outer leaves are adapted to intense light conditions, whereas the shaded interior leaves are larger and thinner.
Interior canopy leaves are loaded with photon-capturing pigments to catch the scraps of light missed by the outer canopy. These interior leaves tend to be horizontally oriented and can be fairly long-lived. Shade leaves are important contributors to photosynthesis and should not be routinely pruned out as a means of tidying up a tree or shrub. To do so injures the plant’s ability to nourish its interior canopy.

Hosta leaves are broad and thin to capture maximum solar energy.

Hostas in full sun can collect too much energy and bleach out.

Dusty miller leaves have dense fuzzy mats for sun protection.
Speaking of pruning, we now have a good idea of why the neighbor’s boxwoods have turned white. The interior leaves, once nestled in deep shade, were suddenly exposed to intense levels of full sun. The pigment-rich tissues absorbed much more sunlight than could possibly be funneled into the photosynthetic apparatus, and that extra energy has nowhere to go. It overloads the chloroplasts and literally fries them from the inside out, a process called photo-oxidative bleaching. Just as if they’ve been dipped in Clorox, the leaves are a pure white. All is not lost, however. Once the bleached leaves fall, new leaves will emerge that are adapted to full sun conditions. Remember this in your own garden, and save pruning for early spring or late autumn when sun injury is less likely.
Looking for That Place in the Sun
On the way back home, we automatically duck under the dogwood tree that leans out into the street. It started out straight enough when it was planted ten years ago, but has progressively grown away from the arborvitae hedge looming behind it. There are all kinds of tree bondage kits available for straightening up recalcitrant trees. Would one be worth a try?
True sun-loving species are garden divas: they are happiest in the middle of the solar spotlight. Surrounded by abundant sunlight, they capture only a fraction of the available radiation for photosynthesis. If moved off center stage into shaded conditions, they never reach their full potential and can languish for years waiting to be rediscovered by the sun.
It’s easy to recognize these unhappy performers, especially when compared to their sun-drenched counterparts. Shaded sun-lovers have sparse leaves that are larger and paler than normal. The stem tissues between the leaves (the internodes) are longer as the plant puts extra energy into outgrowing the shade. Most obvious is the leaning growth habit that they will develop in an effort to reach the sun. The tilt is the result of an uneven growth pattern in which auxins cause the shadiest side of the plant to grow the fastest. The difference in growth rates causes the tree to bend away from the shade. Most of the leaves will be found on the least shaded side of the plant as well, emphasizing the asymmetrical shape.
But plant bondage kits won’t help straighten unhappy sun-lovers. The plants will continue to reach for the sun. The only cure is prevention: put them on center stage far away from those who might steal the spotlight. Also consider using light-colored mulches and other reflective surfaces to add some extra brightness.

Sun-loving species try to escape the constant shading of conifers.
Photosynthetic Parasites
Most of the plants we cherish in our homes and gardens follow the typical photosynthetic lifestyle. But some do not. Thumb through any wildflower guide and you’ll find a small section on plants that aren’t green but instead sport white, brown, or yellow hues. These species are parasitic because they siphon away sugars from hard-working photosynthesizers without giving anything in return. If you were able to burrow underground and follow their roots, you’d find these sponging off the roots of green plants, usually with the assistance of some fungal partners.

Pine drops are the flowering portion of an underground parasitic plant.

Indian pipe emerges through the forest duff layer.
Even though their botanical habits may be unsavory, these parasitic plants are both unusual and beautiful. Indian pipe (Monotropa uniflora) and pine drops (Pterospora andromedea) are just two of these lovely species. Unlike their photosynthetic cousins, these plants are pure white and chocolate brown, respectively. And woe to anyone who illegally digs these plants to use in their own garden, for without their photosynthetic hosts and fungal go-betweens, these species will never survive.
Photosynthetic variants are unusual. We understand the importance of being green. But sometimes normally green leaves transform to the chromatic opposite of green: red. What’s the trigger for this switch?