How Plants Work (2015)
1 Under the Microscope
IT’S TAKEN NEARLY FIFTEEN YEARS, but our garden is finally something I’m happy to share with visitors. In the front yard, a lovely pond dominates the sunny landscape, surrounded by small trees, shrubs, and groundcovers in every imaginable foliar shade: black mondo grass, lime green ginkgo, brick red laceleaf maple, and a contorted larch that turns the most amazing fiery colors in autumn. Following the flagstone pathway around the house and through a narrow glade filled with rhododendrons and azaleas, we enter the shady, north-facing backyard with its fragrantly flowered sarcococca and hardy gardenias. Ferns and mosses, ancient plants, are at home in this environment, as are the hostas with their dinner-plate-sized leaves. Hydrangeas thrive, untouched by pests or disease. On the deck in a sunny spot is my bog garden, a large ceramic container full of carnivorous pitcher plants and sundews.
The curious gardener might wonder why there are so many different leaf colors. What makes those flowers smell so good? How can a tree grow in bizarre contortions and maintain that shape? And why do some plants successfully avoid pests and disease? To answer these and many other questions that gardeners have asked for centuries, we need to step out of the garden and into the laboratory to explore the world of plant cell biology.
This chapter gives you a quick and selective introduction to plant anatomy and biochemistry. There are some structures, chemicals, and processes associated with plant cells that we need to discuss before we can look at how a plant works in its environment. First, we’ll look at several cell structures: some that are found in both plants and animals and others that are unique to plants. Next, we’ll consider the vast pharmacy of biochemicals manufactured by plants to fabricate, communicate, and regulate. Finally, we’ll look at three specialized cells and tissues whose activities we can help—or hurt—when we’re gardening.
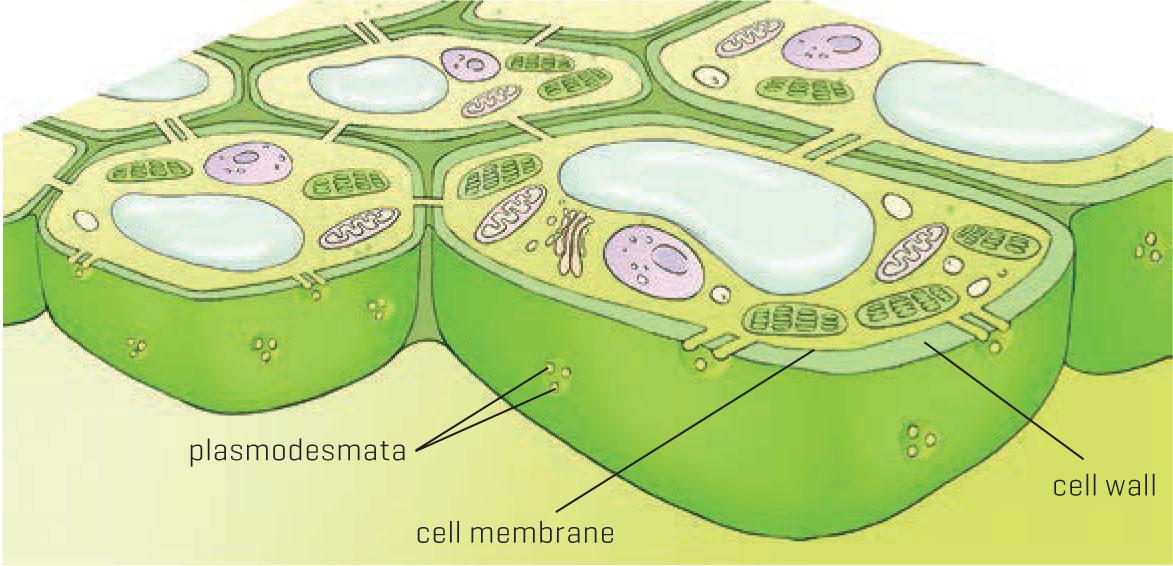
Plant cells are connected to one another by plasmodesmata.
The Cell Wall: Home Protection
Unlike animal cells, plant cells are surrounded by a rigid wall made of cellulose and other biochemicals. Rigid cell walls ultimately allow trees to grow to great heights and protect individual cells from environmental injury and biological attack. But while the cell wall protects the cell and gives it rigidity, it also isolates cells from one another, which makes transport of materials between cells very difficult. Fortunately, the walls are pierced by tiny holes through which the inner cell membranes extend. These tiny membrane tubes, called plasmodesmata, are an important means of transport and communication between cells. Plasmodesma (the singular form) has a very cool Greek meaning: plasm refers to form and desma is a girl’s name meaning binding oath. So plasmodesmata bind forms together, in this case plant cells.
Individual cells are also cemented to one another by a gelatinous layer of pectins, carbohydrates that you’re familiar with if you make preserves. Without pectin, jams and jellies would be more like syrup, as would young plant tissues.
Why do plants have cell walls, rather than some kind of internal skeleton? Plant cells have to withstand internal water pressure, and without the wall, the cell membrane would burst. This water pressure is vital in moving sugars, minerals, gases, and other dissolved substances throughout the plant.
The Cell Membrane: Border Patrol
Both plant and animal cells are surrounded by a membrane, which keeps all the contents floating together in a soupy liquid called cytoplasm. The cells have various organelles (literally, little organs) that have specific tasks, such as producing energy (mitochondria), directing cell reproduction (nucleus), or packaging materials (Golgi apparatus and Golgi bodies). Nutrients, dissolved gases such as oxygen and carbon dioxide, and building blocks for various compounds are all contained in the cytoplasm. Besides keeping all the organelles and cytoplasm contained, the cell membrane plays a critical role in determining what’s allowed to enter and leave the cell.
As an analogy, let’s consider the border between any two countries. I travel frequently between Washington State and British Columbia, and when I enter either country a customs official questions me about fruit, alcohol, and other materials I might be transporting. Likewise, receptors on the cell membrane inspect various molecules to determine whether they should be allowed into or out of the cell. Usually this system works well, but occasionally something unwanted might slip across, like a virus or an environmental pollutant.
The Mitochondria: Power Plants
Those of you who remember your basic high school biology might recall this nifty alliteration: “the mighty mitochondria are the powerhouses of the cell.” All plant and animal cells contain mitochondria. Here’s the interesting thing: the mitochondria are, in fact, alien invaders. Mitochondria contain DNA that’s radically different than the DNA in the cell’s nucleus, and they reproduce independently of the cell’s reproductive cycle. (Fun fact: the mitochondria in most plants and animals are derived from those in the egg. That’s how maternal lines are established.) Mitochondrial DNA is most similar to that found in present-day bacteria.
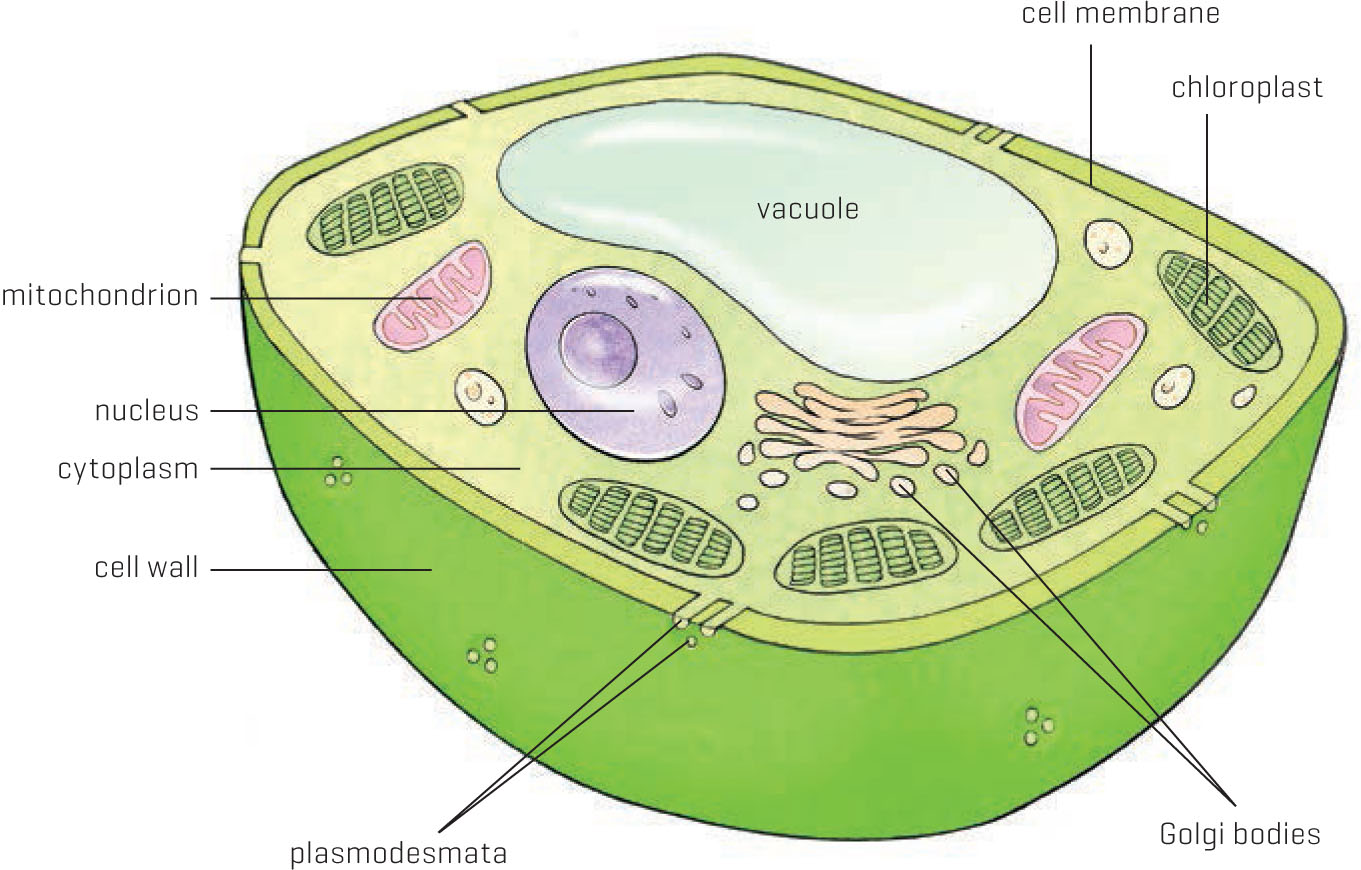
A typical plant cell
How could this possibly have happened? The most convincing theory, which is supported by a great deal of evidence, is that ancient, single-celled ancestors of plants and animals engulfed ancient bacterial cells. This isn’t as far-fetched as you might think. Going back to high school biology again, you might remember watching a paramecium or amoeba (single-celled animals) gobble up little algal or bacterial cells under a microscope. The theory states that some of these bacterial cells were not digested and instead became permanent cellular guests. Because they benefitted the host cell by providing energy, the descendants of the original host cells retained the bacteria, which eventually lost the ability to live independently and are now cell organelles.
The Chloroplasts: More Alien Invaders
One of the obvious differences between plants and animals is that plants photosynthesize using chlorophyll-rich chloroplasts to capture light energy and convert it to sugar. Like mitochondria, chloroplasts contain unique DNA and replicate independently. However, chloroplast DNA is most similar to that found in blue-green algae. So plant cells contain two different alien invaders, both of which benefit the host cell by producing energy.
When plant cells eventually divide, the nucleus orchestrates the action while mitochondria and chloroplasts simply go along for the ride. It’s not an equal partitioning, so dividing cells usually have different numbers of chloroplasts in each half. From a gardener’s perspective, this is where some really interesting things can happen in plants.
Just like nuclear DNA, chloroplast DNA also collects mutations (spontaneous, random changes in the genetic code). Some of these mutations affect the color of the chloroplast itself. If a newly dividing cell only contains white chloroplasts, for instance, then that cell will be white, as will all of its descendants. What started as a single white cell becomes a white blotch and eventually a variegated leaf.
The Vacuole: Warehouse Central
When we look at a plant cell under the microscope, most of it appears to be made up of nothing. That nothing is the vacuole (meaning vacuum). Because plants don’t have an excretory system like animals do, for many years vacuoles were thought to be cellular garbage dumps. The vacuole is a membrane filled with water and lots of dissolved substances: acids, sugars, and an amazing assortment of what are collectively called secondary compounds. In a healthy cell, the vacuole is so bloated that it may take up 95 percent of the entire cell, pressing all the other organelles against the cell membrane. Oddly, this is actually good for cell health, because oxygen, light, and other necessities enter the cell through the membrane.
I don’t like crowds, and I really don’t like crowds in small spaces, like elevators. When I’m in a hot and stuffy crowded elevator, I try to stand along a wall, especially near the front where fresh air rushes in when the doors open. So, although it might seem problematic that the chloroplasts, mitochondria, and other organelles are squashed against the cell membrane by the vacuole, it’s the best possible place for them to be to get oxygen, light, and other resources.
The vacuole is able to absorb and retain all of this water because it stores a lot of dissolved substances. An important mantra in understanding how plants work is “Water always moves to where water isn’t.” The more compounds the vacuole accumulates, the less space is left for water. This imbalance attracts more water into the vacuole, which creates high turgor pressure inside the cell. When the vacuole simply can’t expand any more—when it squashes into the cell wall—then water flow stops.
What about all of those dissolved compounds that vacuoles supposedly store? Are they really waste products, as once thought? Or do they have more important roles to play?
Secondary Compounds: A Plant’s Personal Pharmacy
Many of the dissolved substances within the vacuole are secondary compounds. This name is unfortunate because it suggests these chemicals are not as important as primary compounds. While primary compounds are those required for plant growth and development (things like sugars, DNA, fats, and proteins), various secondary compounds have defensive, attractive, or as yet unknown properties. So, even if they aren’t required for basic growth, secondary compounds are crucial in a plant’s ability to compete and survive in its environment.
I can still remember the “wow” moment when I learned in my graduate plant biochemistry class that we probably have yet to discover 90 percent of the compounds that plants make. I wouldn’t be surprised if the number was closer to 99 percent. Researchers have done a good job of investigating secondary compounds that have economic benefit, from vanilla to pyrethrin to codeine, but it’s likely that every plant species makes one or more unique compounds. We haven’t even scratched the surface.
Secondary compounds are usually classified based on the biochemical pathway through which they are constructed by the plant (such as the alkaloids, phenolics, and terpenoids). This does little to help gardeners (or anyone else) understand what roles they play in a plant’s life. Rather than turn this into a side trip through Biochemistry Land, let’s look at the major roles secondary compounds play in plant survival.
BUILDING BLOCKS
LIGNIN. Every time you get a splinter, struggle to cut through a tree branch, or marvel at the ability of a spirally trained spruce to maintain its shape, you can thank (or blame) lignin. This complex compound toughens up the walls of mature cells and allows them to resist many environmental stresses, pests, and disease. It is slow to decompose, which gives woody mulch its staying power.
CUTIN. Cutin is part of the cuticle, which is the flexible, waxy layer protecting leaves and other soft tissues above ground. Cutin and other associated biochemicals help keep water in and invaders out of leaves. Its waxy nature is sensitive to soap, however, which is why many herbicides contain a soapy substance that helps breach the cuticle and deliver death to weeds.
SUBERIN. While leaves are covered in cutin, roots are swathed in a suberin sheath that has the same protective function. Additionally, suberized cells help select what substances in the roots will be transported through the xylem into the rest of the plant. The suberin in bark is what makes bark mulch a pretty poor choice, as the waxiness repels water absorption and movement.
CAROTENOIDS. Like their name suggests, carotenoids are carroty-orange pigments, although they can range in color from pale yellow to nearly red. Though their presence in leaves is masked by chlorophyll, carotenoids are important in scavenging the green light that chlorophyll can’t capture for the photosynthetic machine.
ENVIRONMENTAL AND DISEASE PROTECTION
ANTIOXIDANTS. Many secondary compounds have antioxidant activity, including carotenoids and anthocyanins, the pigments that make plant tissues orange or various shades of red, blue, and purple, respectively. In plant cells, antioxidants absorb and neutralize reactive chemicals that otherwise could damage sensitive membranes. We get at least some of these benefits when we eat pigment-rich plant parts, and it certainly makes me feel virtuous about my fresh raspberry addiction (especially when they’re out of season locally).
SUNSCREEN. Some compounds, like anthocyanins, absorb excess solar energy so leaves aren’t overloaded. Others, like cuticular waxes, reflect sunlight and reduce the heat load. Plants with gray-green or blue-gray foliage are loaded with these waxes. Indoor and greenhouse-grown plants don’t waste energy on these protective agents, however, and they will fry just like my Irish skin does if they’re moved outside into full sun. Tuck them into shaded areas for several days first to let them toughen up.
ANTIMICROBIAL AGENTS. When attacked by fungal pathogens, plants can use existing saponins, soap-like compounds that break down fungal cell walls. Some other chemicals, called phytoalexins, are made only when pathogens attack. Finally, lignin and other structural compounds can be used as cellular reinforcements to wall off the rest of the plant from disease.

Plants from hot environments, like this manzanita, often have gray-green foliage to reflect excess sunlight.

The blue-gray foliage of this palm is adapted to sunny conditions.
HERBIVORE PROTECTION
Many of the secondary compounds produced by plants to fend off herbivores will repel, deter, or kill you as well. When plants are domesticated for human consumption, we breed many of these defensive compounds out of them. That’s why your fruits and veggies are so inviting to insects and other pests: they no longer have their natural abilities to keep predators away.
Harpin
THE PRODUCT
Sold as Messenger, harpin is derived from a protein isolated from fire blight bacteria (Erwinia amylovora).
THE SUPPOSED BENEFITS
Harpin acts like a vaccination to prepare plants to fight environmental stress and disease.
HOW PLANTS RESPOND
When it enters a plant cell, harpin turns on biochemical pathways that produce anti-stress compounds. It’s a process that works great in the laboratory, but has little success with real plants in the landscape. The reason is pretty easy to understand. Remember that cell wall and those layers of waterproofing compounds on the leaf surface? It’s really difficult to force the harpin molecule through the cuticle and cell wall. If harpin can’t get into the cell, it can’t turn on the necessary biochemical pathways. This is a product that’s a good idea, but with limited practical use.

A protein from the bacterium responsible for fire blight helps plants fight disease in the laboratory, but not in the garden.
REPELLANTS. The odor of these compounds drives plant-eating insects and animals away before they can even take a bite. We take advantage of aromatic repellants, like essential oils of clove, cinnamon, and lemon, as natural pesticides. How many times do you see insect damage on mint leaves as you’re picking them to make jelly or juleps? Obviously, like us, other animals are also attracted to rather than repelled by these compounds.
DETERRENTS. The taste of these compounds keeps snacking herbivores from coming back for seconds. These are often acidic, bitter, or astringent: think Mr. Yuk. Tannins, named for their early use in tanning leather, are good examples of astringent deterrents. When you take a mouthful of a nice dry red wine or bite into a too-green banana, you are getting a dose of tannins. While the first example may be a more pleasant experience than the second, the effect in your mouth is the same: the tannins bind to your salivary proteins and make your mouth feel drier. This doesn’t hurt us, but insects that feed on tannin-rich leaves may have so many of their salivary and digestive proteins bound up that they aren’t able to get much food value from the leaves. Too much of a tannin-rich diet may prevent them from surviving and reproducing. Successful insects look for something a little less astringent.
Other deterrents include skin irritants, like the itchy oils of poison ivy, poison oak, and poison sumac, and phototoxic chemicals found in many members of the parsley family, such as giant hogweed. Farm workers and even grocery shoppers have gotten itchy hands from these light-activated chemicals when handling damaged or diseased celery or parsnips.
POISONS. Of course, these chemicals are the ultimate deterrents. Murder mystery fans will be familiar with strychnine (an alkaloid found in seeds of an Asian tree) and digitalin (a cardiac disruptor found in foxglove leaves). Other less infamous poisons can interfere with growth and reproduction of herbivorous insects. It’s not surprising, then, that many of these compounds, like pyrethrins, neem, rotenone, and nicotine, are widely used as pesticides. (But just because these are natural pesticides, it doesn’t mean they are safer than synthetic ones.)
ATTRACTION
Plants not only produce secondary compounds to repel other organisms, they also emit chemical signals into the environment to attract them. For example, beneficial microbes—bacteria and fungi that provide food, protection, or other benefits—often rely on chemical cues from receptive plants as their invitation to begin a partnership.
Sending out an SOS
Like clockwork, the first buds on the rose bushes in our sunny front yard emerge in April, followed by an army of aphids that covers the buds entirely. If I happen to see this, I’ll set my hand sprinkler on stun and blast them away, but sometimes I’ll forget. When that happens, do my rosebuds get sucked away into lifeless husks? No. In fact, they don’t show much damage at all. Eventually our local lacewings and ladybugs stop by for a little green snack.
How do these beneficial predators know where the aphids are? Many plants, ornamentals and vegetables alike, send out very specific gaseous signals when they’re under attack. Over time, certain species of predatory and parasitoid insects have learned that these airborne alarms mean lunch. These signals are only emitted during the day, when natural enemies are active. When the herbivorous pests are gone (having either escaped or been eaten), the compounds are no longer produced. To make this phenomenon even more fascinating, plants downwind of the victim may also pick up on the signal and start building chemical defenses against future attack by the herbivore.
When we gardeners indiscriminately spray pesticides for the slightest pest problem, we not only kill the pest, but also the beneficial organisms that could take care of our problem for us at no charge.
POLLINATION. Plants are beguiling in the scents and colors they use to attract birds, bees, and a whole slew of other animal pollinators. Essential oils and other volatile compounds lure pollinators with promises of food or even sex. Your nose might even have helped in pollination as you moved through your garden, taking in all the fragrances. Flower colors are provided by secondary compounds and can be quite specific in their targets. Birds, for instance, see best in the red end of the spectrum—hummingbirds are an obvious example. In contrast, bees prefer flowers at the blue end of the spectrum and can even see into the ultraviolet range. Though we can’t see them, many flowers take advantage of bee vision and provide pollination guides leading to the center of the flower that can only be seen under ultraviolet light. If you were a bee, you’d pretty much be seeing giant neon signs reading “Eat here!”
DISPERSAL. After successful pollination, scents and colors are used to attract animals as seed dispersers. The volatile compounds associated with ripe strawberries, peaches, and other summer delights, heated and released by the sun, are irresistible to us and the other fruit eaters we race against. Colors, provided by anthocyanins, carotenoids, and other pigments, are important visual cues that alert fruit lovers to edibility. Whereas most green fruits are hard and bitter, their more mature red, blue, or orange forms are magnets for attention.
GROWTH
ALLELOPATHIC CHEMICALS. Because plants cannot move around to find the resources they need, they can be fierce competitors, exploiting resources and sometimes interfering with their neighbor’s ability to do the same. Allelopathic chemicals (meaning killer of others) seep from roots, leaves, fruit, and/or bark of some species, endlessly frustrating gardeners who can’t figure out why nothing will grow under their walnut, eucalyptus, or tree-of-heaven. Gardeners have taken advantage of some of these allelopathic chemicals to use as natural herbicides.
PHYTOHORMONES. Some secondary compounds either stimulate known plant growth substances or take this role on for themselves. You’ll notice I said “plant growth substances” and may wonder why I’m so stuffy that I can’t just say “plant hormones” like other people. The main reason is that it causes confusion. People naturally assume that plant hormones behave like hormones in animals. In reality, they don’t. In general, phytohormones can be produced by all plant cells, in contrast to animal hormones that are produced in glands. So, I’ll compromise with the term phytohormones, as long as you remember that they are very different from those swarming around in your own body.
Phytohormones can stimulate or inhibit plant activities and control everything from growth to reproduction to death. We’ll consider some specific phytohormones later, but it’s worth introducing the main groups.
Vitamin B1
THE PRODUCT
Vitamin B1 (often mixed with fertilizers or plant hormones) is added to new transplants.
THE SUPPOSED BENEFITS
Vitamin B1 stimulates new root development and reduces transplant shock.
HOW PLANTS RESPOND
Plants make their own vitamin B1, so adding it does absolutely nothing for the plant. However, those formulations that include plant hormones such as auxin can help root development. If you are installing picky plants or are working in difficult soils, you could try a rooting hormone. But don’t bother with these mixed products, especially those that contain phosphate fertilizer.

Cytokinins produced by insects that lay their eggs within leaf tissue often create galls as nurseries.

Uncontrolled growth in this conifer has created a witches’ broom at the top.
Auxins are phytohormones that control rooting, stem elongation, and directional growth. Charles Darwin was the first to propose the existence of this group. Gardeners know about auxins because they’re used as rooting powders that cuttings are dipped into. As we’ll discover later, they’re also important in controlling branching. Actively growing shoot tips crank out auxins, which are shuttled downward to other parts of the plant. Auxins are also instrumental in directional growth, causing shoots to grow toward light and roots to grow downward.
Cytokinins are involved in cell division or cytokinesis. Many are manufactured in the roots and transported to shoots, but they’re also found in young leaves and fruits, tissues that are rapidly growing. I’ve referred to them at times as fountain-of-youth phytohormomes, because they can help delay the aging of leaves. Gardeners are familiar with burls, galls, and witches’ brooms, which are often caused by foreign cytokinins made by bacteria, fungi, viruses, nematodes, and even insects. The supercharged growth rate of the infected host provides a constant source of nutrition and protection for the invader—a botanical room and board as it were. This relationship can sometimes benefit the plant, too: nitrogen-fixing bacteria create nodules and mycorrhizal fungi develop filamentous hyphae on roots through this same process.
Gibberellins regulate plant height and flower, fruit, and seed development. The first gibberellin was identified in rice plants in Japan suffering from foolish seedling disease. This wonderfully named disease causes rice seedlings to grow so tall that they fall over. It also reduces the affected plant’s ability to set seed. Gibberellins can be thought of as the maturing phytohormones—kind of the opposite of cytokinins. As you might suspect, dwarf cultivars of many plants don’t synthesize much gibberellin, and adding gibberellins to dwarf cultivars allows them to grow normally. The commercial use of gibberellins has been valuable in increasing fruit size, especially of seedless cultivars like grapes. G is for giant grapes and gibberellins!
Abscisic acid was named for its presumed role in abscission, the deliberate shedding of leaves, fruits, and other plant tissues by programmed cell death. Abscisic acid is particularly active during times of environmental stress, especially any stress associated with lack of available water. In fact, abscisic acid can directly reduce water loss by closing off the leaf pores (stomata), tiny portals through which gases and water vapor travel. Abscisic acid also regulates seed and bud dormancy. Generally, it’s considered to be an inhibitory phytohormone, as it tends to slow growth rather than enhance it.
Ethylene is a gas. Really! Put green tomatoes into a bag with a ripe banana, and ethylene released from the banana seems to magically transform rock-hard tomatoes into soft and juicy delights. Ethylene is also produced by various stresses, so it teams up with abscisic acid in plant responses such as leaf drop, wound sealing, and disease resistance. Like abscisic acid, ethylene tends to be an inhibitory phytohormone associated with programmed tissue death. If you perpetually overwater a container plant, eventually you will see the results of the ethylene gas produced by drowning roots: the lower leaves turn yellow and fall off, leaving only the leaves on top of a bare stem.

Overwatered plants often drop their lower leaves due to the release of ethylene from stressed roots.
Brassinosteroids are a tough-sounding group of phytohormones. They are chemically related to testosterone and other steroids, but you don’t have to worry about your marigolds getting muscle-bound. Brassinosteroids are synthesized throughout the plant and are involved in all aspects of growth and development, both stimulatory and inhibitory. They’ve been used commercially to improve crop yield in plants growing under stressful conditions and can be used like auxins to root cuttings.
Biostimulants
THE PRODUCT
Also called metabolic enhancers, biostimulants include humic acids, kelp, probiotics, and other low-nutrient products.
THE SUPPOSED BENEFITS
Biostimulants improve various plant responses including growth, yield, and stress resistance, possibly through stimulation by plant hormones found in the product.
HOW PLANTS RESPOND
Plants make their own phytohormones, and an intact plant is unlikely to benefit from additional hormones applied to the soil or leaves. While there is some value to using hormone dips to root cuttings, there is no consistent positive effect of biostimulants used in any other manner. Proper plant selection, installation, and management practices that ensure development of a healthy root system are more likely to improve growth, yield, and stress resistance than biostimulants. In fact, that’s what research has shown.
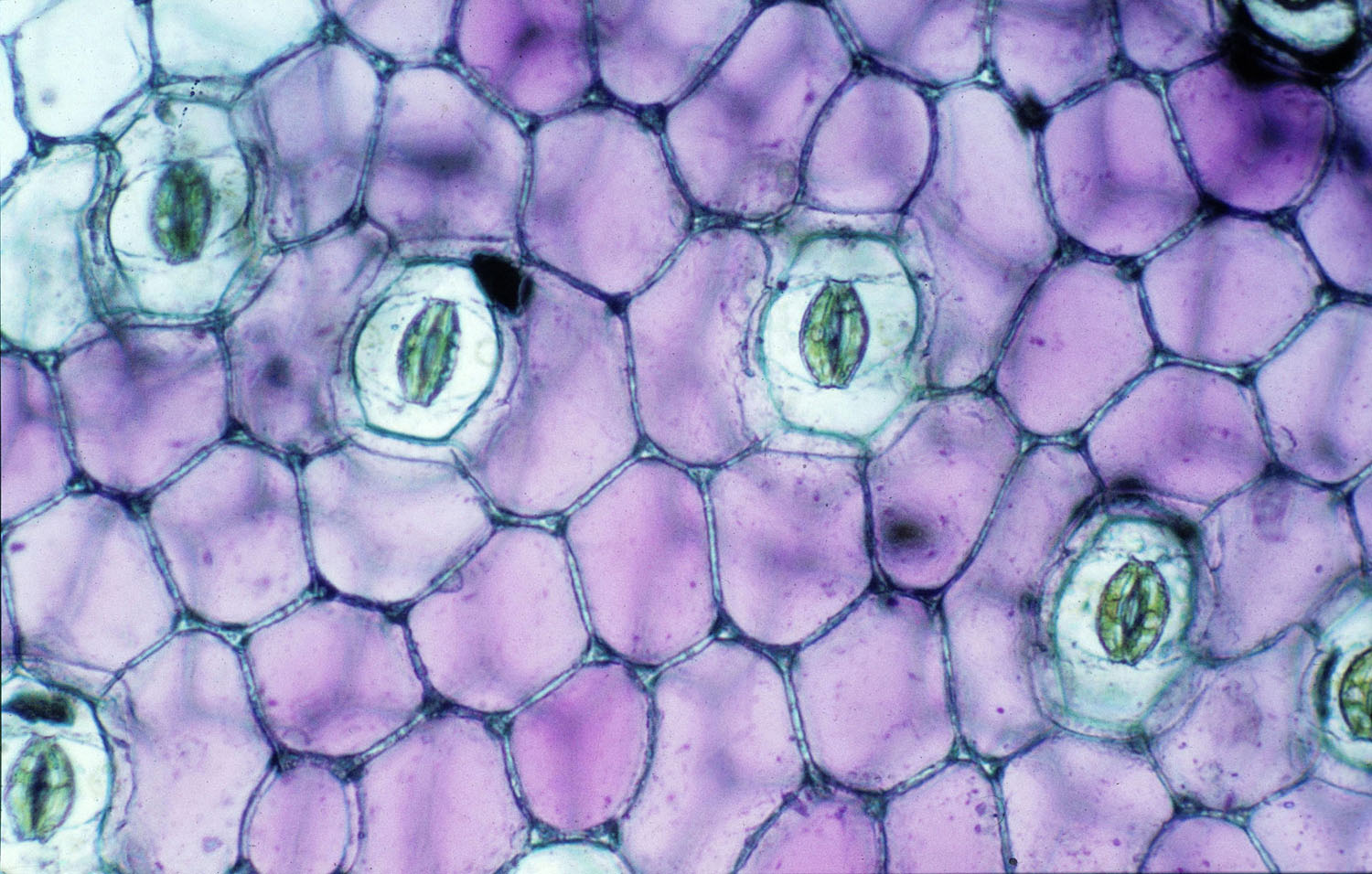
Leaf stomata consist of pores bordered by two guard cells.
Putting It All Together
We’ve looked at some of the unique qualities of plant cells, which help us understand—and sometimes predict—how plants will respond to their environment. We’ll be using this information in later chapters. Let’s look at a few nifty plant tissues under the microscope to get us started.
STOMATA
If you were to look at the lower surface of a hydrangea leaf from my garden using a strong hand lens, you would see stomata (also called stomates), pores scattered like tiny pouty lips over the surface. Stomata are the leaves’ gas and water regulators. Whereas people have an active respiratory system that brings in the good air and lets out the bad, oxygen and carbon dioxide move passively through these stomata. The pores are also the exit doors for water moving through the plant.
Simple stomata have a pore bounded by two kidney-shaped guard cells, which are attached to one another at their ends. When the guard cells are full of water, their sides push away from each other, opening the pore to the environment. When the leaves wilt, the guard cells are flaccid and the edges seal over the pore, thus reducing water loss.
You’ll want to remember how this works later when we look at environmental conditions that cause stomata to open and close and how that affects plants.
APICAL AND LATERAL MERISTEMS
We’re back with the microscope, this time looking at clusters of visually unremarkable cells at the tips of roots and shoots. It’s at these points, the apical meristems, where cells are rapidly dividing and elongating, allowing roots and shoots to lengthen. Beyond this region, mature plant cells become specialized for water uptake, storage, or some other function. Growing points are also contained in buds tucked into the junctions between leaf petioles and stems; under the right conditions, these buds will develop into branches or flowers. And one group of plants, the grasses, grows from the base rather than the tips.
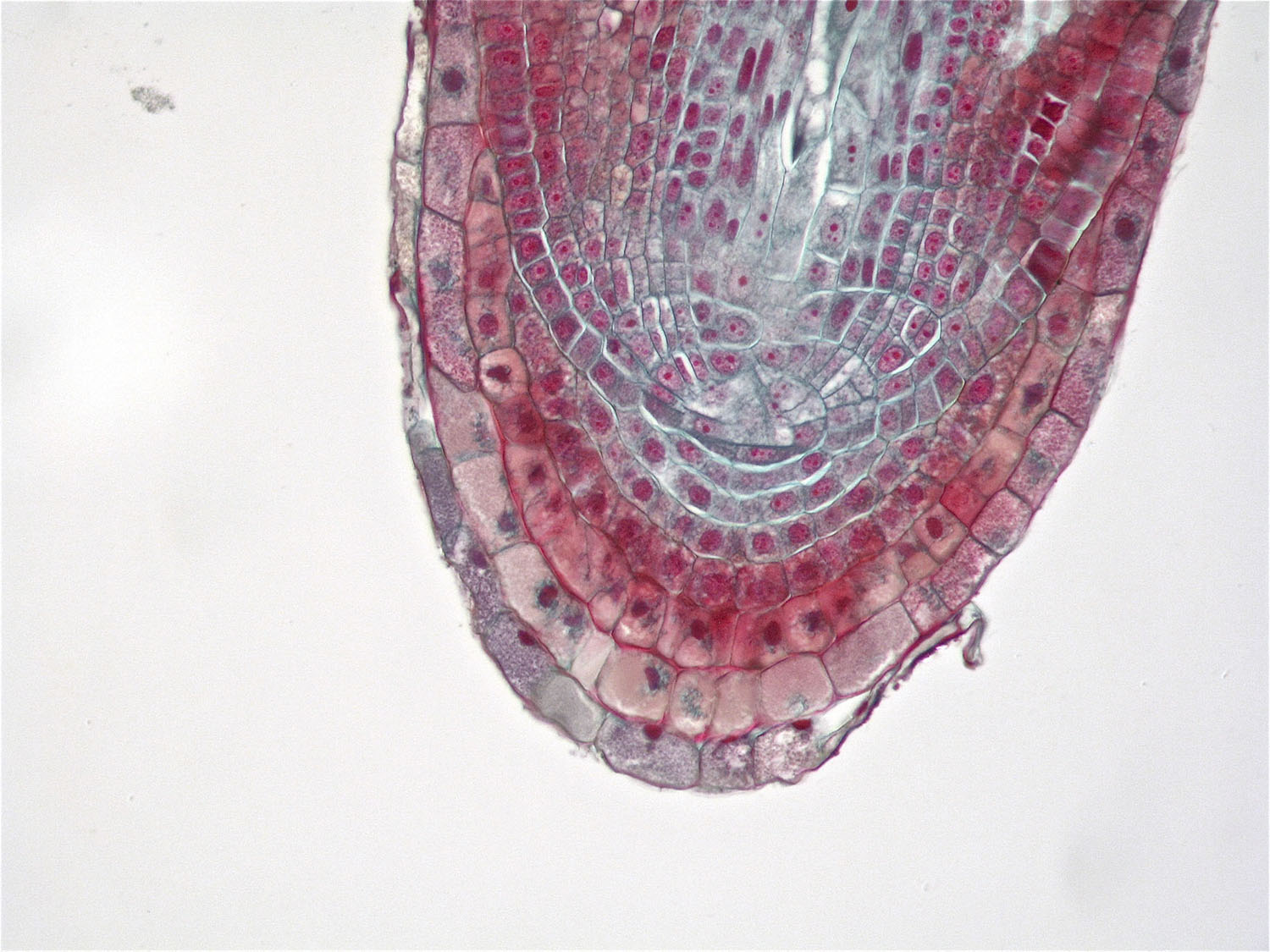
When mature, these rapidly dividing young root tip cells will have specific functions.
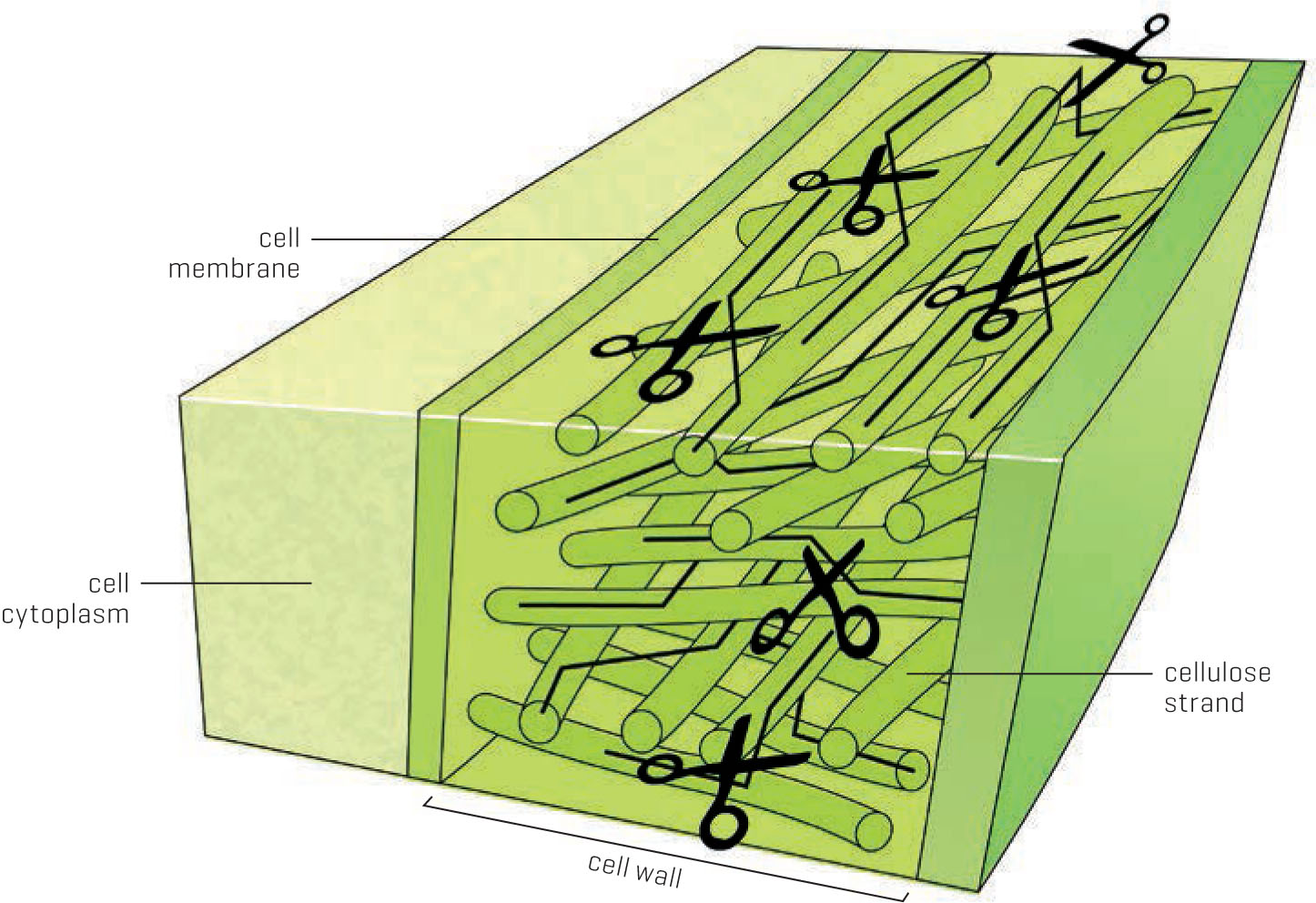
The cell wall is composed of a latticework of cellulose strands. When the crosslinkages are severed, the strands can slip by each other, allowing the plant cell to elongate.
Apical meristems allow stems and roots to lengthen, whereas other areas of growth increase the girth of branches, trunks, and roots. These are the lateral meristems, found just under the outer protective layers of bark. These lateral meristems include the vascular cambium, which produces new conducting elements (xylem and phloem), and the cork cambium, which produces bark-related tissues. Lateral meristems are found only in perennial plants that develop girth with age.
“But wait!” the curious gardener will exclaim. “You said that cell walls are rigid. How can they elongate?” We’ll start with an easy demonstration to visualize this. Intertwine your fingers together, squeeze them together tightly, and then try to pull them apart. Now relax your fingers and slide them apart without separating them completely. The elongation process in the cell walls is very similar. The cellulose strands in the wall become looser and slipperier during the growth phase, like strands of spaghetti, allowing the cell to lengthen. Once this window of opportunity closes, the strands are locked into place and the cell has formally reached its maximum size.
It’s fun to step back from the microscope at this point and consider some analogies. The meristematic cells, undeveloped and actively growing, are like children. As growth slows and they mature, cells take on separate identities and functions, much like teenagers and young adults choose career paths. And of course, the development of girth is something seen in older plants and people!
Understanding where plant growth can and cannot take place will help you understand how plants respond to gardening activities like pruning and staking, as well as how you can do it correctly.
We’ve had a very quick and selective introduction to plant anatomy and biochemistry. Congratulations on slogging all the way through! You’ll find it was well worth your time and patience in subsequent chapters. And I guarantee you will revisit this chapter, both as you continue exploring this book and your own garden.
In any case, I hope you don’t feel you’ve been left in the dark. That’s a condition best suited for roots, which may be out of sight, but they certainly shouldn’t be out of mind.